- Published:
Kimmeridgian protoglobigerinids (Foraminifera) from Crussol (SE France)
Swiss Journal of Geosciences volume 103, pages 83–100 (2010)
Abstract
A detailed investigation of the protoglobigerinids from the Kimmeridgian section of the Montagne de Crussol, SE France is presented. This monotonous succession of predominantly carbonate beds represents peri-platform hemipelagic deposits, well dated by ammonites. Three species were determined in thin sections and in isolated forms after extraction by acetolysis. Favusella? parva (Kuznetsova) is the most common species occurring in almost the entire succession from the Orthosphinctes to the Beckeri zones. Favusella hoterivica (Subbotina) has been retrieved less frequently from the Platynota to the Beckeri zones, and it is its oldest occurrence. The rare Globuligerina bathoniana (Pazdrowa) occurs only in the Late Kimmeridgian. Based on our study and the review of the literature a palaeogeographical map was plotted, showing that most of the protoglobigerinid occurrences are in deep water facies of the northern margin and central part of the Tethys. These records demonstrate the presence of Globuligerina bathoniana, G. oxfordiana (Grigelis), Favusella hoterivica, F.? parva and Compactogerina stellapolaris (Grigelis) in the Kimmeridgian.
Introduction
Protoglobigerinids are tiny globigerina-like foraminifera known from the Jurassic sediments, especially from the Oxfordian for a long time. They are mentioned from the Late Triassic to the Early Cretaceous in rock thin sections as well as in isolated forms. However, the finds in isolated forms are very scarce, sporadic in space and time. Generally, they are poorly preserved because of their aragonite test wall. Despite the exhaustive synthesis of BouDagher-Fadel et al. (1997) their detailed morphology of these fossils, taxonomy and stratigraphic and palaeobiogeographic distributions, are still very lacunal. To understand the phylogenetic origin of these foraminifera, their evolution and their hypothetic relations with the Cretaceous planktonic species request to improve the field records as well as the laboratory analysis. While the Middle Jurassic–Oxfordian protoglobigerinid record is fairly good, there is a huge gap in the knowledge of this group from the Kimmeridgian up to the Hauterivian (e.g. Riegraf 1987a; BouDagher-Fadel et al. 1997; Görög and Wernli 2003, 2004). In these stages, some protoglobigerinids are reported in rock thin sections, while the descriptions of isolated forms are dramatically scarce.
Hence, the aim of this work is to review the Kimmeridgian records of protoglobigerinids in the literature and to add new discoveries in thin section as well as in isolated forms from the well-dated field section of Crussol, stratotype of the Crussolian stage, near Valence in the south-eastern France (Fig. 1).

Location of the studied Crussol section. Scale bar 50 km
Previous researches on the Kimmeridgian protoglobigerinids
Kimmeridgian protoglobigerinids were identified in 44 publications concerning 26 areas or lithologic units (Table 1; Fig. 2). The Kimmeridgian age of these fossils is questionable in 11 papers, and in 14 occurrences, the age is Kimmeridgian without any further specification. In most cases (17) the authors mentioned the forms from the Early Kimmeridgian and only three times from the Late Kimmeridgian. From 10 areas (21 papers) only citations without illustrations could be found. The protoglobigerinids are figured in thin sections of 10 areas (14 works) and as isolated specimens from 6 areas only (9 papers). Based on our experience (Görög and Wernli 2003) high and low trochospiral forms could be identified in the published illustrations and descriptions, suggesting that the low trochospiral forms are more frequent.

Palaeogeographical distribution of the Kimmeridgian protoglobigerinids. The palaeogeographical map is modified after Thierry (2000). Numbers refer to the Table 1. ACP Apennine Carbonate Platform, AM Armorican Massif, Aq Aquitaine, BK Bakony, Bu Bükk, CPP Corbières-Provence Platform, Dm Dalmatia, Dr Drama, EP Eastern Pontids, FC Flemish Cap, GrB Great Bank, H Hochstegen, IM Iberian Meseta, LBM London Brabant Massif, M Moesia, Ma Mardin, MC Massif Central, Mg Magura, MM Moroccan Meseta, OMB Oued Mya Basin, RM Rhenish Massif, SC Silesian Cordillera, Sic Sicanids, SP Serbo Pelagonian, TrP Trento Plateau, UMB Umbria-Marches Basin, VoB Vocontian Basin, VB Volgian Basin, WP Western Pontids
Ten species were cited, half of them from only one locality and we could not find any detailed systematic description about Globigerina cf. bulloides d’Orbigny, 1826, ?Globuligerina lobata (Terquem, 1883) and “Globigerina (suprajurassica ?)” (in Bergougnan 1971). In 27 publications, the determinations indicate only Globigerina or “Protoglobigerina”.
In our detailed analysis, we disregard citations without illustration. For the morphological description of the shells, H is the height of the trochospire and D is the largest transverse diameter.
Records in rock thin sections in the literature
Our compilation shows that Kimmeridgian protoglobigerinids in thin sections are sparsely illustrated in the literature (Table 1). Apart from a single location in the North Atlantic (Luterbacher 1972), the palaeogeographical distribution of these occurrences covers essentially the Northern and the Central parts of the Tethyan realm (Fig. 2). Only one record is located in epicontinental sea (Southern Jura Mountains, Bernier 1984).
The determinations are usually on generic level. In general the sections are small, with D being in average around 180–200 μm, sometimes reaching up to 270 μm. In the majority of the areas the high and low trochospiral forms are associated. The walls appear thin, always recrystallised, and were probably originally aragonite. The building of the tests are not very clear, it may be consecutive or concurrent (Wernli 1988).
The most detailed studies are those of Giovagnoli and Schiavinotto (1986, 1987, 1990), who analysed a great number of sections from the Early Kimmeridgian of the Apennines (Umbria-Marche facies). Their statistical results show that the Kimmeridgian forms are small (up to 240 μm) with high trochospire, thin wall and concurrently built test. Hereinafter we will demonstrate that low trochospiral as well as high trochospiral forms exist in the Kimmeridgian.
Records in isolated forms in the literature
Grigelis et al. in (1977, p. 927, fig. 1) described by means of drawings a new, low trochospiral species, Globuligerina stellapolaris (Grigelis, 1977) from the Kimmeridgian and the Volgian (Tithonian) of the Timan-Pechora region in northern Russia. Later, Grigelis and Gorbatchik (1980a, pl. 2, fig. 1, republished in 1980b, pl. 1, fig. 5) and Gorbatchik and Kuznetsova (1998, pl. 1, figs. 1–3) gave SEM pictures of topotypes. This species is also cited from the Early Kimmeridgian of Crimea (Kuznetsova and Uspenskaya 1980; Gorbatchik and Kuznetsova 1998). The diameter is 250–260 μm and the wall surface shows pseudomuricae in discontinuous ridges. Simmons et al. (1997) have chosen G. stellapolaris as type species of their new monotypic genus Compactogerina, and they only recorded it from the Tithonian.
In northwestern Atlantic at DSDP site 534A, the poorly preserved Globuligerina aff. oxfordiana (Grigelis, 1958) (Gradstein, 1983, p. 552, pl. 2, figs. 1–10, low trochospiral small forms, D = 70–80 μm) are probably Oxfordian in age, even though the dinoflagellates indicate Kimmeridgian age.
Kuznetsova and Gorbatchik (1985, p. 113, pl. 16, figs. 1, 2) illustrated by means of drawings their new species Globuligerina parva (Kuznetsova, 1985) from the Early Kimmeridgian of Crimea. The re-illustrations by SEM in Gorbatchik and Kuznetsova (1998, pl. 2, figs. 8, 9) reveal that the holotype and the paratype are poorly preserved.
El Kamar (1986) published pictures with few visible details of Globuligerina oxfordiana (pl. 2, fig. 8, pl. 5, figs. 7, 10) and Globuligerina bathoniana (Pazdrowa, 1969) (pl. 3, figs. 1, 6; pl. 5, fig. 4) from the earliest Kimmeridgian of Portugal.
Stam (1986) took good pictures of different species form the Early Kimmeridgian (Platynota Zone) of Portugal, like Globuligerina oxfordiana (pl. 8, figs. 4–12) (figs. 9–12 could be Favusella? parva), Globuligerina bathoniana (pl. 10, figs. 9–12) and Globuligerina balakhmatovae (Morozova, 1961) (pl. 11, figs. 11, 12), even thought the latter forms could be tectonically deformed specimens of G. oxfordiana.
Samson (1997, p. 142, pl. 16, figs. 12–19) illustrates poorly preserved specimens of Globuligerina oxfordiana from the Early Kimmeridgian of region of Havre, Seine-Maritime, Normandie, France. The individual on plate 16, fig. 12 (re-illustrated in Samson 2001, pl. 5, fig. 30) with a low extraumbilical aperture could belong to Haeuslerina helvetojurassica (Haeusler, 1881). Those attributed to Globuligerina cf. balakhmatovae are deformed (pl. 16, figs. 21–23) while a high trochospiral form (pl. 16, fig. 20) to Globuligerina bathoniana.
Simmons et al. (1997) illustrated by SEM a metatype of Haeuslerina parva (=Favusella? parva). In their synthetic table “Stratigraphical distribution of Conoglobigerinidae” (op. cit., fig. 2.2) only two species (H. parva and G. bathoniana) occur in the Early Kimmeridgian, but not any in the Late Kimmeridgian.
To sum up, the record of the isolated protoglobigerinids in the Kimmeridgian is very poor in the number of sites as well as in the quality of the material. Moreover, the published illustrations do not allow unambiguous identifications. Almost all the finds come from the northern margin and the central part of the Tethys and from the North Atlantic. For these areas, we can retain the occurrences of Globuligerina oxfordiana, G. bathoniana and Favusella? parva. Only Compactogerina stellapolaris has been found in high latitude (Northern Russia).
The geological setting of the Crussol field section
The studied field section is located in the Montagne de Crussol, near Valence (Ardeche Department), Rhone valley, in SE France (Fig. 1). During the Kimmeridgian, the Crussol area was situated in the northwestern edge of the Vocontian Trough, at the European margin of the Tethys, which formed a gulf opening to the east (Fig. 2). In this direction, it grades into the deep facies of the Valais Trough that is connected with the Alpine Tethys Ocean (Thierry 2000). The Vocontian Trough is bounded to the west, south and southeast by the Corbières-Provence shallow carbonate platform. Exposed lands in proximity are the Massif Central to the west and the Corsica-Sardinia block to the southeast. From the Late Oxfordian to the Early Kimmeridgian the Vocontian Trough is subjected to a subsidence inducing an extension and the enlargement of the basin on its margins. From the Late Kimmeridgian the slowdown of the subsidence coupled with a compression phase produced the progradation of the neritic facies toward the centre of the basin (Enay 1984; Colombié and Strasser 2003).
Materials and methods
The Montagne de Crussol is an abrupt rocky mountain with several abandoned quarries on its western flank, which provide a continuous section from the Oxfordian to the Tithonian. The Kimmeridgian part of the succession has been studied in detail by Atrops (1982) who determined the ammonites. He numbered the beds in the outcrops so we could use his numbering in our work (Fig. 3). Langoisseux (2001, unpublished) restudied the same outcrops carrying out a sequence stratigraphy using multidisciplinary approach. His collection of rock thin sections (Cru) has been used for the present paper (Fig. 3).

Lithostratigraphic log of the Crussol section (simplified after Atrops 1982) and distribution of the protoglobigerinids. X no protoglobigerinids, Cru collection of O. Langoisseux, W collection of R. Wernli
The Kimmeridgian section can be observed from the base to the top, going upwards on the mountain, in the quarry D, then in the quarry B and finally in the quarry F following the nomenclature of Atrops (1982, fig. 55). From the Panula Zone (partly) to the Beckeri Zone (partly) the sampled succession measures about 70 m in thickness.
This is a monotonous series of decimetric to plurimetric limestone or marly limestone beds generally without interbeds, with plane or wavy beddings. Only rare centimetric or thicker marl interbeds can be found. The limestones are essentially mudstones, micritic, beige, grey or pink-grey in colour, with few grains visible in the field. The outcrop represents a typical peri-platform, hemipelagic deposit. Apart from the ammonites, the other macrofossils (belemnites, brachiopoda, aptychii, echinoderms) are relatively rare. The microfossil content is analysed below in the chapter “Micropalaeontological associations”.
From the Kimmeridgian succession, four marl samples have been washed by standard method but we failed to find any protoglobigerinids in these facies. In calcareous indurated rocks, 41 thin sections of rocks collected by O. Langoisseux (coll. Cru) and eight by us (coll. W) have been analysed. All these samples are deposited in the Département de Géologie et Paléontologie, Université de Genève. Eleven samples of calcareous and marly rocks have been treated by means of acetolysis following the method of Lethiers and Crasquin-Soleau (1988). This extraction gave better results than the standard washings. Our records of protoglobigerinids in thin sections and in isolated forms are given in Fig. 3.
Micropalaeontological associations of the Crussol section
In rock thin sections the microfossils are relatively rare, forming a very monotonous and constant association in these mudstones all along the succession. There are tuberoids (sensu Gaillard 1983), small Globochaete, recrystallized radiolarians (dominant spumellarians), calcispherulids (Colomisphaera spp.), Saccocomidae, microgastropods, microbrachiopods and some “filaments” and aptychii. The foraminifera fauna is subordinate: besides the protoglobigerinids, it contains small, simple agglutinated forms (Ammobaculites irregularis (Gümbel, 1862), Reophax sp.), tiny Nubeculariidae, some Nodosariidae (Lenticulina, Nodosaria), Polymorphinidae and Spirillinidae.
The record of the isolated microfossils extracted by standard washing as well as by the acetolysis is very variable from bed to bed. Hence, the plankton–benthos ratio (P/P + B) varies from 5 to 40%. However, these figures must be used with caution because of the inconsistent extraction method. This technical distortion can be well illustrated in some samples, for example at the base of the outcrop (W2617), where the protoglobigerinids are present in rock thin sections but absent in the acetolysis residues. Hereinafter we give only a succinct analysis because the benthic foraminifera have not been studied in detail.
In the Early Kimmeridgian the associations are poor and less diverse, with radiolarians, some Nodosariidae (mostly Lenticulina), Spirillina, Polymorphinidae and some agglutinated species and Paalzowella. The Late Kimmeridgian reveals a richer and more diverse fauna: dominant groups are lenticulinids (Lenticulina muensteri Roemer, 1839, L. subalata (Reuss, 1854), Astacolus varians (Bornemann, 1854), L. quenstedti (Gümbel, 1862), Polymorphinidae (Eoguttulina oolithica (Terquem, 1874), E. bilocularis (Terquem, 1864), Guttulina pera Lalicker, 1950, Sigmomorphina ssp.) and spirillinids (Spirillina elongata Bielecka Pozaryski, 1954, S. tenuissima Gümbel, 1862). Other Nodosariidae (Dentalina and Nodosaria) are relatively diverse, but not frequent; smooth and ornamented forms are also present. In some samples agglutinated specimens are also abundant, mostly small-sized Haplophragmoides, and Glomospira. Besides them, rhizamminids, Textularia jurassica Gümbel, 1862, Ammodiscus sp., Reophax sp. and Ammobaculites sp. also occur. The epistominids are rare.
All samples of the succession contain radiolarians, sponge spicules, saccocomids, other echinoderm fragments, microgastropods, ammonite protoconchs and a few ostracods.
In all samples, the abundance of the protoglobigerinids diminishes when the number of radiolarians increases.
Protoglobigerinids of the Crussol section
Protoglobigerinids in rock thin sections
Almost all 49 rock thin sections analysed from Crussol yield protoglobigerinids (see Fig. 3). They are scarce, usually about ten per slide (1–2 sections by cm2) and never appearing as bloom like in the oceanic Middle Jurassic. The most frequent sections are small, about 70–160 μm in the largest diameter, some rare reaching up to 250 μm. A large part of them are entirely filled with sparicalcite. Some are partially filled, generally in the early whorls, leaving only the last chambers empty. In this case the thickness of the wall measures about 6–7 μm, which is always recrystallised in sparicalcite, that is also visible on some isolated forms (Fig. 4.6). No sections clearly show the details of the juvenile stage. In several sections displaying the three last successive empty chambers, the thickness of the wall seems to be constant and so the building of the test appears consecutive.

Scale bars are 200 μm in 1–2, 4–5, 7–9 and 11–14, and 10 μm in 3, 6, 10 and 15. umb. umbilical view, lat. lateral view, enlarg. enlargement. 1–11Globuligerina bathoniana (Pazdrowa), 1 lat.-umb. W2619, 2 lat. W2764, 3 enlarg. of 2, 4 umb. W2766, 5 umb. W2766, 6 enlarg. of 5, 7 umb. W2768, 8 umb. W2769, 9 umb. W2769, 10 enlarg. of 9, 11 lat.-dorsal, W2769. 12–15Favusella? parva (Kuznetsova), 12 umb. W2765, 13 lat. W2766, 14 umb. W2769, 15 enlarg. of 14
The specific, even generic determination of the protoglobigerinids based on sections is very difficult (Wernli and Görög 2000). In the case of the Kimmeridgian of Crussol where there are isolated shells as well as sections from the same levels, it is possible to tentatively attribute:
-
the high trochospiral forms to Globuligerina cf. bathoniana (Fig. 5.7, 5.9, 5.12, 5.14),
Fig. 5 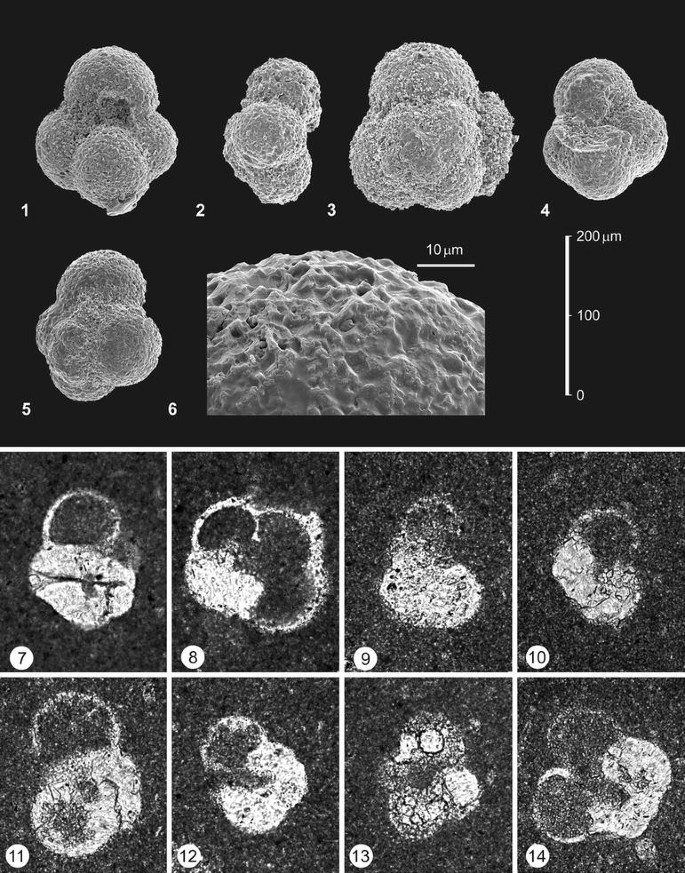
Scale bars are 200 μm in 1–5 and 7–14, and 10 μm in 6. umb. umbilical view, lat. lateral view, spiral spiral view, enlarg. enlargement. 1–6Favusella hoterivica (Subbotina), 1 umb. W2619, 2 lat. W2768, 3 spiral with brocken bulla, W2763, 4 umb. with brocken bulla, W2769, 5 spiral oblique, W2619. 7–14 Protoglobigerinids in rock thin sections. The numbers indicate the bed numbers. 7, 9, 12, 14Globuligerina cf. bathoniana (Pazdrowa), 7, 8 156, 9, 12 119, 14 W2768. 8, 10 undetermined protoglobigerinids. 8 156, 10 W2768. 11Favusella cf. hoterivica (Subbotina), W2768. 13Favusella? cf. parva (Kuznetsova), W2766
-
the low trochospiral forms to Favusella cf. hoterivica (Fig. 5.11),
-
the small petaloids equatorial sections to Favusella? cf. parva (Fig. 5.13).

The habit of the protoglobigerinids of Crussol well coincides with those of Giovagnoli and Schiavinotto (1990) from the Kimmeridgian of Apennines. In our opinion, the building of the tests (consecutive or concurrent) cannot clearly be identified in both occurrences. We can confirm contrary to these authors (op. cit., fig. 12) and also Wernli (1988), that high trochospiral tests as well as low trochospiral ones exist together in the Kimmeridgian (Figs. 3, 5.7–5.14).
Protoglobigerinids in isolated forms
In general, the isolated tests are moderately well preserved and not deformed. Although the wall is recrystallized in some cases the surface ornamentation is still retained. The number of the specimens strongly varies from sample to sample, from 3 to 5 up to 150. The vertical distribution of the different species can be seen in Fig. 3.
Superfamily Favusellacea Longoria, 1974 emend. Banner and Desai, 1988
Family Conoglobigerinidae Simmons, BouDagher-Fadel, Banner and Whittaker, 1997
Genus Globuligerina Bignot & Guyader, 1971, emend. Simmons, BouDagher-Fadel, Banner and Whittaker, 1997
Globuligerina bathoniana (Pazdrowa, 1969)
Figure 4.1–11.
- ?1961:
-
Globigerina conica n. sp.; Iovčeva & Trifonova, p. 343, pl. II, figs. 1–8.
- pars 1969:
-
Globigerina bathoniana n. sp; Pazdrowa, p. 45, pl. I, figs. 1, 2, 4 (non fig. 3), figs. 5–9, pl. II, figs. 1–5, 7?, 8, 9, pl. III, fig. pl. IV, 1–3. non pl. II, figs. 6 ab.
- ?1973:
-
Conoglobigerina bathoniana (Pazdrowa, 1969); Fuchs, p. 455, pl. 1, fig. 2 (glauconitic moulds).
- 1976:
-
Gubkinella bathoniana Pazdro (sic); Ascoli, p. 672, 713, pl. 1, fig. 1.
- pars 1976:
-
“Globigerina” bathoniana (Pazdrowa); Gradstein, p. 560, pl. 3, fig. 1 non? 2ab. (transitional form to G. oxfordiana).
- 1977:
-
Globigerina hoterivica Subbotina; Masters, p. 460, pl. 22, figs. 1–3.
- 1980a:
-
Conoglobigerina bathoniana (Pazdro) (sic); Grigelis & Gorbatchik, p. 9, pl. 1, figs. 3a–c. Refigured in:
-
1980b Conoglobigerina bathoniana (Pazdro) (sic); Grigelis & Gorbatchik, pl. 1, figs. 3ab (topotype).
-
1981 Conoglobigerina bathoniana (Pazdro) (sic); Alekseeva & Gorbatchik, pl. 1, figs. 1ab.
- 1981 ?:
-
Globigerina bathoniana Pazdro (sic), 1969; Bielecka & Styk, p. 34, pl. 3, figs. 21ab.
- pars1986:
-
Globuligerina bathoniana (Pazdrowa); Stam, p. 108, pl. 9, figs. 6, 7, 9, 11–13; pl. 10, figs. 1–12; pl. 11, figs. 1–10; pl. 13, figs. 1–12; pl. 14, figs. 1, 2, 5–7. (non pl. 9, figs. 8, 10). The pl. 9, fig. 12 is refigured in:
-
1988 Globuligerina bathoniana (Pazdrowa); Banner & Desai, pl. 1, fig. 4.
- 1987a:
-
Globuligerina bathoniana (Pazdrowa, 1969); Riegraf, p. 195, pl. 1, figs. 1–11 and text-figs. 5a–e (drawings). Reillustrated in:
-
1987b Globuligerina bathoniana (Pazdrowa); Riegraf, figs. 3–13.
-
1988 Globuligerina bathoniana (Pazdrowa); Riegraf, p. 474, pl. 5, figs. 23–25.
- non 1996:
-
Globuligerina bathoniana (Pazdro) (sic); Kuznetsova et al., p. 191, pl. 15, fig. 14.
- 1997:
-
Globuligerina bathoniana (Pazdrowa, 1969); Simmons et al., p. 27, pl. 2.8, figs. 4–12.
- 2002:
-
Globuligerina bathoniana Pazdrowa (sic); Görög & Wernli, p. 30, pl. 1, figs. 28, 29.
Description: The relatively small tests reaching the maximum 200 μm in the largest dimension are high trochospiral (H/D > 0.9) with \( 3 \frac{1}{2} \text{--} 4\) chambers in the last whorl. The general shape is roughly conical with a rounded apex giving a U profile in lateral view. The juvenile stage has not been clearly analysed because the chambers are strongly appressed. On the umbilical side, the last \( 3 \frac{1}{2} \text{--} 4\) chambers are globular with incised sutures giving a strongly lobulated outline. The aperture is an umbilical, asymmetric, globuligerine-like arch as high as wide surrounded by a lip. The distal part of this arch (following the ontogenic progression) is protruding when the proximal one is re-entrant into the chamber (Fig. 4.4). The aperture is commonly damaged, in this case it appears like a single arch without lip. The umbilicus is well marked, depressed, but in the majority of the specimens it is filled by sediment therefore the aperture is not clearly visible.
The wall is generally recrystallised with a surface showing the quadrate and rhombic network of the diagenetic sparicalcite (Fig. 4.6). Only in some case, we can see a surface with few elevated, more or less vermiculated discontinuous pustules (Fig. 4.10) but never forming a meshwork like in the favusellids. Perforations have not been observed.
In the rock thin sections the wall measures about 7 μm in thickness and the building of the tests seems to be consecutive.
Remarks:Globigerina conica Iovčeva & Trifonova from the Tithonian of Bulgaria is a high trochospiral form strongly resembling to G. bathoniana in general shape. However, all specimens are glauconitic molds, without characteristic details of the wall and the aperture.
In the original description, Pazdrowa (1969) accepts a very large variability for her new species G. bathoniana especially in the height of the trochospire. However, we consider the middle to high trochospiral forms with a minimum H/D ratio of 0.9 as G. bathoniana, and the low trochospiral forms as Globuligerina oxfordiana (Grigelis) (pl. II, fig. 6ab). This applies also to Gradstein’s (1976, pl. 3, fig. 2ab) and Stam’s specimens (1986, pl. 9. figs. 8, 10). Disregarding the height of the trochospire G. bathoniana is strongly related to G. oxfordiana as illustrated by of Pazdrowa (1969), Stam (1986) or Simmons et al. (1997).
The specimens of Kuznetsova et al. (1996) have an exceptional large size, 380–460 μm, and very low trochospires.
Occurrence:G. bathoniana occurs in the Late Kimmeridgian of the Crussol section, relatively in small number.
Distribution: Late Bajocian—Late Kimmeridgian (Tithonian?).
Ogrodzieniec area, north from Krakow, Poland: Bathonian (Pazdrowa 1969; Masters 1977; Grigelis and Gorbatchik 1980a, b; Alekseeva and Gorbatchik 1981; Stam 1986, Simmons et al. 1997), Late Callovian–Early Oxfordian (Fuchs 1973); North Western Poland: Bathonian (Bielecka and Styk 1981); Scotian shelf: Bathonian?–Callovian (Ascoli 1976); Grand Banks, New Foundland: Late Bathonian–Early Callovian (Gradstein 1976), Late Bajocian–Bathonian (Stam 1986); Brenha section, Portugal: Early Bathonian (Stam 1986); Mareta Beach, Portugal: Callovian (Stam 1986); Tojeira 1 section, Portugal: Early Kimmeridgian (Platynota Zone) (Stam 1986); SW Germany: Callovian (Riegraf 1987a, b, 1988); Bakony Mts, Hungary: Bathonian (Görög and Wernli 2002); Stubel village, Moësian Platform, NW Bulgaria: Tithonian (Iovčeva and Trifonova 1961).
Family Favusellidae Longorina, 1974
Genus Favusella Michael, 1973
Favusella hoterivica (S ubbotina , 1953 )
- 1953:
-
Globigerina hoterivica n. sp.; Subbotina, p. 50, pl. 1, figs. 1–4. fide Catalogue Ellis and Messina. Fossil foraminifera of the USSR: Globigerinidae, Hantkeninidae and Globorotaliidae. VNIGRI 76, Leningrad.
- ?1961:
-
Globigerina terquemi n. sp.; Iovčeva & Trifonova, p. 344, pl. II, figs. 9–14.
- 1964:
-
Globigerina tardita Antonova sp. n.; Antonova et al., p. 60, pl. 12, figs. 4a–c, 5a–c.
- 1964:
-
Globigerina quadricamerata Antonova sp. n.; Antonova et al., p. 60, pl. 12, figs. 6a–c.
- 1976:
-
Caucasella hoterivica (Subbotina); Ascoli, p. 674, pl. 1, fig. 3.
- 1980a:
-
Globuligerina hauterivica (sic) (Subbotina); Grigelis & Gorbatchik, pl. 2, figs. 3ab.
- 1980b:
-
Globuligerina hauterivica (sic) (Subbotina); Grigelis & Gorbatchik, pl. 2, figs. 7ab.
- 1981:
-
Globuligerina hauterivica (sic) (Subbotina) 1953; Gorbatchik & Antonova, p. 84, pl. 1, figs. 1–4. Refigured in:
-
1986 Globuligerina hauterivica (sic) (Subbotina); Gorbatchik, pl. 11, figs. 1ab, 2.
-
1988 Favusella hoterivica (Subbotina); Banner & Desai, pl. 1, figs. 6ab.
- 1986:
-
Globuligerina quadricamerata (Antonova); Gorbatchik, pl. 12, figs. 1a–c, 2ab, 3. The pl. 12 figs. 1bc refigured as:
-
1988 Favusella tardita quadricamerata (Antonova); Banner & Desai, pl. 1, figs. 8ab.
- 1986:
-
Globuligerina tardita (Antonova); Gorbatchik, pl. 13, figs. 1 ab, 2, 3. Refigured in:
-
1988 Favusella tardita (Antonova); Banner & Desai, pl. 1, figs. 7ab.
- 1988:
-
Globuligerina hoterivica (Subbotina); Ascoli, pl. 1, figs. 14–16.
- 1995:
-
Favusella hoterivica (Subbotina); Wernli et al., p. 388, pl. 1, figs. 1–7, pl. 2, figs. 1–16, pl. 3, figs. 1–15; Text-fig. 5. The pl. 4.2, figs. 1–8 refigured in Simmons et al. 1997.
- 1997:
-
Favusella hoterivica (Subbotina, 1953); BouDagher-Fadel et al., p. 60, pl. 2.5, figs. 11–14 (same pictures on pl. 4.1. figs. 2–5!), pl. 4.1, figs. 1, 6–17, pl. 4.2, figs. 1–8.
- ?2004:
-
“transitional forms between Globuligerina oxfordiana (Grigelis, 1958) and Favusella hoterivica (Subbotina, 1953)”; Görög and Wernli, p. 41, pl. 1, figs. 3–11.
Description: The small tests between 180 and 220 μm in the largest diameter are low trochospiral (H/D = 0.6–0.8) with typically four chambers, sometimes 3½ in the last whorl. These last chambers increasing regularly in size are fully globular and separated by incised sutures giving a well quadrilobated umbilical side. The aperture is (intra-) umbilical in position drawing a weakly asymmetric arch, as wide as high, surrounding by a lip. Its form is more regular and less “globuligerine” than in Globuligerina oxfordiana and there is no fossa.
On the spiral side which is moderately convex we count 11 or more chambers, since the details of the first whorls are not visible, coiled in at least three whorls. The arrangement of the chambers is relatively compact and the juvenile part is blunted, not forming a protruding apex.
A large part of the material is recrystallised and shows the quadrate and rhombic texture of the shell, typical of the diagenetic calcite. Only in some cases, there appears a reticulate surface ornamentation with small irregular and polygonal meshes of 5–7 μm in diameter that is diagnostic of the genus Favusella (Fig. 5.6). No perforations have been observed.
Some of the shells have a bulla variable in size that can be flat, small or as large and inflated as the last chamber. Its position is more or less umbilical. On a broken isolated individual the thickness of the wall of the bulla is about 10 μm (Fig. 5.4).
In rock thin sections the wall is about 7 μm in thickness and the building of the tests seems to be consecutive.
Remarks: Distinguishing F. hoterivica from G. oxfordiana can be difficult on poorly preserved tests without arguments regarding the wall, and if the aperture is not clear. Besides, transitional specimens between the two species probably occur in the Kimmeridgian–Tithonian (Görög and Wernli 2004). Following our experience:
-
F.hoterivica has a larger, more symmetrical, less virguline aperture than G.oxfordiana.
-
F.hoterivica has no fossa. In G. oxfordiana the position of the aperture is not strictly umbilical but slightly shifted toward the distal direction. The re-entrant (proximal) part of the aperture produces a narrow umbilical depression (fossa). This feature is well developed in the Oxfordian individuals of G. oxfordiana (see for example illustrations in Bignot and Guyader 1971; Wernli and Kindler 1986; Grigelis and Norling 1999; Simmons et al. 1997).
-
F.hoterivica more frequently possess a bulla. A big bulla can be confused with a normal last chamber and raises the question of the standard orientation of the test as discussed by Wernli et al. (1995). The bulla is not situated in the line of the trochospiral ontogenic progression. It also differs from the rest of the test in its whitish colour and less hyaline, sugar-like aspect. It never shows a reticulate or favose surface.
-
F.hoterivica generally has a more compact test with more fully globular chambers.
Occurrence: F. hoterivica occurs in the Early as well as in the Late Kimmeridgian of the Crussol section, relatively frequent.
Distribution: Early Kimmeridgian–Early Aptian.
Pshish, NW Caucasus: Hauterivian (Subbotina 1953; Gorbatchik 1986; Gorbatchik and Antonova 1981); Shise and Tusheps river, Mazutka, NW Caucasus: Late Barremian–Early Aptian (Antonova et al. 1964; Gorbatchik 1986; BouDagher-Fadel et al. 1997); Kura river, NW Caucasus: Early Aptian (Antonova et al. 1964; Gorbatchik 1986; BouDagher-Fadel et al. 1997); 397A site, N Atlantic: Late Barremian (Gorbatchik 1986); NW Atlantic margin of North America: Valanginian (Ascoli 1988); Scotian shelf: Valanginian (Ascoli 1976), Berriasian–Valanginian (Wernli et al. 1995); Urukh river, N. Caucasus: Hauterivian (Grigelis and Gorbatchik 1980a, b).
Favusella ? parva (K uznetsova , 1985)
- non 1973:
-
Globuligerina parva n. sp.; Fuchs, p. 466, pl. 4, fig. 6.
- ?1983:
-
Globuligerina aff. oxfordiana (Grigelis); Gradstein, pl. 2, figs. 9, 10. (surface with tubercules). ? pl. 2, figs. 1–8 (strongly corroded specimens).
- 1985:
-
Globuligerina parva K. Kuznetsova, sp. nov.; Kuznetsova (in Kuznetsova and Gorbatchik), p. 113, pl. 16, figs. 1a–c, 2ab. (drawings).
- 1986:
-
Globuligerina oxfordiana (Grigelis); Stam, pl. 8, figs. 9–12; pl. 14, figs. 13–15.
- ?1997:
-
Haeuslerina parva (Kuznetsova, 1985); Simmons et al., p. 29, pl. 2.5, fig. 4. (badly preserved).
- 1998:
-
Tenuigerina parva (K. Kuznetsova, 1985); Gorbatchik and Kuznetsova, pl. 2, figs. 8a–c (re-illustration of the holotype by MEB), and fig. 9, paratype.
Description: The tests are very small, rarely reaching 150 μm, the majority being between 100 and 120 μm in larger diameter. The trochospire is very low, the H/D ratio is about 0.6. The four well globular chambers in the last whorl give a petaloid outline in umbilical view. The aperture is intra-umbilical in position, forming a regular arch as wide as high, underlined by a lip. The flattened spiral side has not been clearly analysed because of the compact arrangement of the juvenile stage. The wall surface that is rarely preserved sometimes shows a weak polygonal reticulation (Fig. 4.15) similar to that of F. hoterivica, which explains the questionable placement of our individuals in the genus Favusella.
Remarks: The holotype of Kuznetsova (in Kuznetsova and Gorbatchik 1985) is illustrated by a schematic drawing not showing clearly the aperture. The author writes that it is umbilical, in small arch bordered by a weak lip. In 1998, Gorbatchik and Kuznetsova re-illustrated the holotype by means of SEM, but the pictures did not reveal any additional information. An enlargement of the test surface (pl. 2, fig. 8c) shows, according to these authors, low tubercules and pores, which in our opinion are not convincing. The paratype (fig. 9) is strongly corroded and deformed.
The metatype illustrated with the SEM by Simmons et al. (1997) has the umbilical area strongly damaged, does not give any information about the form and the position of the aperture. These authors include parva in the genus Haeuslerina considering that the aperture is intra-extraumbilical.
Despite the inadequate definition of Favusella? parva, that has been little cited and illustrated in the literature, we tentatively attribute our individuals of Crussol to this species.
Occurrence:Favusella? parva is the most frequent forms in our material, occurs in all the Kimmeridgian of Crussol.
Distribution: Late Oxfordian–Late Kimmeridgian.
Meganom Peninsula, East Crimea: Early Kimmeridgian (Kuznetsova and Gorbatchik 1985; Gorbatchik and Kuznetsova 1998; Simmons et al. 1997); DSDP site 534, Blake-Bahama Basin; Oxfordian–Kimmeridgian? (Gradstein 1983); Tojeira section, Mareta beach, Portugal: Late Oxfordian–Early Kimmeridgian (Stam 1986).
Results and comparisons
For study of the protoglobigerinids of the classical Kimmeridgian outcrop of Crussol, 49 rock thin sections, 11 acetolysis extractions on hard rocks and four standard washings in the rare marl interbeds has been analysed. In all rock thin sections the microfossils are rare. The protoglobigerinids are also scarce and never appear as oozes. Their distribution is constant in all the succession. Generally speaking, the determination of the Jurassic protoglobigerinids from sections is very difficult (Wernli and Görög 2000), but in the present work they could be compared with the isolated forms from the same levels. Thus, the high trochospiral sections can tentatively be attributed to Globuligerina cf. bathoniana (Pazdrowa), the low to middle trochospiral ones to Favusella cf. hoterivica (Subbotina) and the small sized, low trochospiral forms to F.? cf. parva (Kuznetsova). These results demonstrate that high and low trochospiral shells coexist in the Kimmeridgian contrary to our previous opinion (Wernli 1988) and to that of Giovagnoli and Schiavinotto (1990). In all thin sections the wall is recrystallized in sparicalcite and the building of the tests seems to be consecutive.
All isolated protoglobigerinids come from hard rock treated by acetolysis. Their abundance as well as the plankton–benthos ratio is very variable due to the facies of the rocks and to the other factors. For example, the aragonite lysocline could play an important role in the preservation of the shells. It well coincides that the abundance of the protoglobigerinids diminishes when the amount of the radiolarians (indicating deeper environment) increase. Generally speaking, in this type of sequences the acetolysis on hard rocks gives better results than the standard washing of marly interbeds, indicating probably a lower sedimentation rate of these calcareous beds.
The isolated protoglobigerinds of the Crussol section are moderately preserved and undeformed, but the apertures are frequently obscured. The wall, which was originally aragonitic is recrystallized in calcisparite and only in rare cases retains the original ornamentation of its surface.
The associations of the isolated protoglobigerinids of the Kimmeridgian of Crussol comprise the following three species:
-
High trochospiral Globuligerina bathoniana, with typical globuligerine aperture and wall with vermiculated discontinuous pustules.
-
Low to middle trochospiral Favusella hoterivica with wall showing the characteristic favusellid ornamentation. Contrary to Globuligerina oxfordiana, it has no umbilical fossa and more frequently possesses a bulla.
-
Small, low trochospiral F.? parva reveals a reticulated surface ornamentation, justifying its transfer from the genus Globuligerina into the genus Favusella.
F.? parva is distributed almost throughout the entire section studied from the Orthosphinctes to the Beckeri zones. F. hoterivica has been retrieved less frequently from the Platynota to the Beckeri zones, when the rarer Globuligerina bathoniana occurs only in the Late Kimmeridgian.
Concerning the isolated forms, the comparisons of our results with the literature is difficult because of the few available data, the poor preservation of the shells and often, the poor quality of many of the published pictures. On the base of the best recognizable illustrations, we can draw the following record of the Kimmeridgian protoglobigerinids:
-
The most common species of the Crussol section, Favusella? parva previously has been known only from Crimea (Kuznetsova and Gorbatchik 1985; Gorbatchik and Kuznetsova 1998).
-
Globuligerina bathoniana is reported of the Early Kimmeridgian of Portugal (El Kamar 1986; Stam 1986) and France (Samson 1997), and appears in the Late Kimmeridgian of the Crussol section.
-
Occurrences of Globuligerina balakhmatovae (Morozova) in the Kimmeridgian are highly doubtful because of all the figured specimens in the literature look like deformed tests of G. oxfordiana or G. hoterivica. According to Wernli and Görög (2007), the stratigraphical range of this species is Late Bajocian–Late Bathonian.
-
Globuligerina oxfordiana and affinis was in offshore NW Atlantic (Gradstein 1983) and Portugal (El Kamar 1986; Stam 1986; Samson 1997).
-
Compactogerina stellapolaris (Grigelis) is known from the Early Kimmeridgian of Crimea and from the Kimmeridgian–Tithonian of Pechora basin, N Russia.
In summary, the protoglobigerinid associations of the Crussol section differ essentially from those of other Tethyan areas, by the absence of Globuligerina oxfordiana and the presence of Favusella hoterivica. Furthermore, the latter shows here its oldest occurrence.
Regarding the Late Jurassic records of the protoglobigerinids worldwide (e.g. Gorbachik and Kuznetsova 1983; Gorbatchik and Kuznetsova 1998; Simmons et al. 1997; present work) we can draw the following provisional scheme of the stratigraphical range of the different species:
-
Globuligerina bathoniana is known from the Early Bajocian (Wernli and Görög 1999), continues into the Oxfordian and becomes extinct in the Kimmeridgian–Tithonian. The status of the similar high trochospiral Globigerina conica Iovčeva & Trifonova (Tithonian) is actually not clear because it is based on glauconitic moulds. It is known only from the type locality and could be a synonym of G. bathoniana. We note that no high trochospiral forms are known in the Berriasian–Valanginian.
-
F. hoterivica has already appeared in the Kimmeridgian, continues into the Tithonian (Görög and Wernli 2003), Berriasian and Valanginian (Wernli et al. 1995) and up to the Early Aptian (BouDagher-Fadel et al. 1997, fig. 4.1). It evolves slowly from G. oxfordiana by simplification of the aperture and acquisition of a favusellid network. Favusella? parva is recorded in the Early and Late Kimmeridgian.
-
Compactogerina stellapolaris (Kimmeridgian–Tithonian) seems to be the only species appearing in high latitude.
-
Neither turborotalid nor hedbergellid forms with extra-umbilical aperture like Haeuslerina helvetojurassica (Haeusler) have been found in post-Oxfordian deposits in the Tethyan realm.
Conclusions
The Kimmeridgian of the Montagne Crussol, SE France is represented by a monotonous succession of predominantly hemipelagic, micritic limestones. The section well dated by ammonites has been analysed in detail by means of numerous rock thin sections, acetolysis extractions and standard washings.
The relatively well-preserved associations of the isolated protoglobigerinids comprise three species: Globuligerina bathoniana, Favusella hoterivica and F.? parva. The smallest form F.? parva (based on surface ornamentation it was classified to genus Favusella) is the most common species distributed almost throughout the entire section from the Orthosphinctes to the Beckeri zones. Globuligerina hoterivica was identified less frequently from the Platynota to the Beckeri zones, while the rarer Globuligerina bathoniana occurs only in the Late Kimmeridgian. Similar faunas have been recognised in the rock thin sections.
The exhaustive review of the literature reveals that the records of the Kimmeridgian protoglobigerinids are rare and generally of poor quality.
The protoglobigerinid associations of the studied Crussol section essentially differ from that of the other Tethyan faunas by the absence of G. oxfordiana and the presence of F. hoterivica, showing its oldest occurrence at Crussol.
Based on our study we can amend the synthesis of Simmons et al. (1997) and BouDagher-Fadel et al. (1997), table fig. 4.1.) about the Kimmeridgian protoglobigerinids:
-
In the Early Kimmeridgian, besides the F.? parva and G. bathoniana, F. hoterivica, G. oxfordiana and C. stellapolaris also exist,
-
We demonstrate the occurrence of F.? parva and F. hoterivica in the Late Kimmeridgian.
Data plotted on a palaeogeographical map show that the majority of the hitherto known occurrences come from the northern margin and central part of the Tethys besides some from the North Atlantic.
Hence, the data of the present study add very important contribution to the knowledge of the Kimmeridgian protoglobigerinids and fill an important gap in the stratigraphical range, the palaeogeographical distribution and the evolution of these foraminifers in the Late Jurassic.
References
Alekseeva, L. V., & Gorbatchik, T. N. (1981). On morphology and systematization of foraminifera analysed by electron microscope. (The use of electromicroscopic analysis in studying the morphology and systematics of foraminifers.). Akademia Nauk SSSR, Voprosy micropaleontologii, 24, 88–94.
Antonova, Z. A., Schmygina, T. A., Gnedina, A. G. & Kalugina, O. M. (1964). Neocomian and Aptian Foraminifera from the Pshekha-Ubin interfluve (northwestern Caucasus) [in Russian]. In: Egoian, V. L (Ed.), Stratigraphy and lithology of the Mesozoic and Cainozoic deposits of the Krasnodarsk area. Trudy Krasnodarskii Filial Vsesoyuznogo Neftegazonogo Nauchno-Isledovatelskogo Instituta 12. 3–72.
Ascoli, P. (1976). Foraminiferal and ostracod biostratigraphy of the Mesozoic-Cenozoic, Scotian Shelf, Atlantic Canada. Maritime Sediments. Special Publication 1 In: First international symposium on benthonic foraminifera of continental margins, part B: paleoecology and biostratigraphy, pp. 653–771.
Ascoli, P. (1988). Berriasian and Valanginian foraminiferal zonation of the NW Atlantic Margin of North America. Benthos’ 86, Revue de Paléobiologie Vol. Spec. 2, 271–280.
Atrops, F. (1982). La sous-famille des Ataxioceratinae dans le Kimméridgien inférieur du S-E de la France. Systematique, évolution, chronostratigraphie des genres Orthosphinctes et Ataxioceras. Documents des Laboratoires de Géologie Lyon 83, 463 p.
Azema, J., Foucault, A., Fourcade, E., Garcia-Hernandez, M., Gonzalez-Donoso, J. M., Linares, A., Linares, D., Lopez-Garrido, A. C., Rivas, P. & Vera, J. A. (1979). Las microfacies del Jurásico y Cretácico de las Zonas Externas de las Cordilleras Béticas, 83 p.
Banner, F. T., & Desai, D. (1988). A review and revision of the Jurassic-Early Cretaceous Globigerinina, with especial reference to the Aptian assemblages of Speeton (North Yorkshire, England). Journal of Micropalaeontology, 7, 143–185.
Barwicz-Piskorz, W., & Tarkowski, R. (1984). Foraminifer assemblages and stratigraphy of Upper Jurassic in Aleksandrów, Near Lód. Bulletin of the Polish Academy of Sciences. Earth Sciences, 32, 81–89.
Beaudoin, B. (1967). A propos de la répartition des Globigérines au Jurassique supérieur et au Crétacé inférieur. Comptes Rendus Academie Sciences Paris, 264(D), 446–449.
Beaudoin, B. (1972). Contribution à l’application des méthodes de l’analyse sédimentaire, a la reconstitution d’un bassin de sédimentation. Exemple du Jurassique terminal—Berriasien des Chaînes Subalpines méridionales. Unpublished PhD thesis, University of Caen, 144 p.
Beaudoin, B. (1977) Méthodes d’analyse sédimentaire et reconstitution du bassin: Le Jurassique terminal—Berriasien des Chaînes Subalpines méridionales. Unpublished PhD thesis, University of Caen, 339 p.
Bergougnan, H. (1971) Remarques stratigraphiques et paléontologiques sur le Jurassique supérieur de la région de Morez (Jura Central). Colloque du Jurassique, Luxembourg 1967. Mémoires de Bureau des Recherches Géologiques et Minieres 75, 249–261.
Bernier, P. (1984). Les formations carbonatées du Kimméridgien et du Portlandien dans le Jura méridional Documents des Laboratoires de Géologie de Lyon, Stratigraphie, micropaléontologie. Sédimentologie, 92(2), 445–803.
Bernoulli, D., & Renz, O. (1970). Jurassic carbonate facies and new ammonite faunas from western Greece. Eclogae geologicae Helvetiae, 63, 573–607.
Bielecka, W., & Styk, O. (1981). Biostratigraphy of the Bathonian and Callovian deposits of North-Western Poland based on occurrence of foraminifers and ostracods. Prace Instytutu Geologicznego, 100, 56.
Bignot, G. & Guyader, J. (1971). Observations nouvelles sur Globigerina oxfordiana (Grigelis). In: Proceedings of II. Planktonic conference, Edizioni Tecnoscienza, Roma, 79–83.
Borza, K. (1959). Geologicko-petrografické pomery mezozoika Belanskych Tatier a massivu Širokej. Geologický Sborník Slovenska Akademia Ved, 10, 133–177.
Borza, K. (1969). Die Mikrofazies und Mikrofossilien des Oberjura und der Unterkreide der Klippenzone der Westkarpaten. Verlag der Slowakischen Akademie der Wissenschaften, Bratislava, 301 p.
Borza, K. (1980). Litologicko-mikrofaciálna charakteristika vrchnojurských a spodnokriedových sedimentov belianskej série (Strážovské vrchy), Lithological-microfacial characteristic of Upper Jurassic and Lower Cretaceous sediments of Belá Group (Strážovské vrchy Mountains). Geologické práce, Správy, 74, 33–56.
BouDagher-Fadel, M.K., Banner, F.T. & Whittaker, J.E. (1997). The early evolutionary history of planktonic foraminifera. London: Chapman & Hall, 269 p.
Colom, G., & Rangheard, Y. (1966). Les couches a protoglobigérines de l’Oxfordien supérieur de l’ile d’Ibiza et leurs équivalents a Majorque et dans le domaine Subbétique. Revue de Micropaléontologie, 9, 29–36.
Colombié, C., & Strasser, A. (2003). Depositional sequences in the Kimmeridgian of the Vocontian Basin (France) controlled by carbonate export from shallow-water platforms. Geobios, 36, 675–683.
Costea, I. & Comşa, D. (1969) Organismes planctoniques a la limite Jurassique-Crétacé dans la Plate-forme moësienne (Roumanie). In: Proceedings of the First International Conference on Planktonic Microfossiles, 1967, Genève, II, pp. 100–122.
d’Orbigny, A. D. (1926). Tableau méthodique de la classe des Céphalopodes. Annales des Sciences Naturelles Paris ser. 1, 7, 96–169.
Dragastan, O., Mutiu, R., & Vinogradow, C. (1975). Les zones micropaléontologiques et la limite Jurassique-Crétacé dans la plate-forme moësienne. Colloque sur la Limite Jurassique-Crétacé, Lyon-Neuchâtel September 1973. Mémoires de Bureau des Recherches Géologiques et Minieres, 86, 188–203.
Dufaure, P. (1958). Contribution à l’étude stratigraphique et micropaléontologique du Jurassique et du Néocomien, de l’Aquitaine a la Provence. Revue de Micropaléontologie, 1, 87–115.
El Kamar, A. (1986). Les protoglobigérines et les microfaunes associées de quelques gisements de l’ Oxfordien de l’Europe occidentale. Unpublished University thesis, University Lyon-1, 152 p.
Ellis, B. F. & Messina, A. (1940). Catalogue of Foraminifera. New York: American Museum of Natural History.
Enay, R. (1984). Jurassique supérieur, paléogéographie. In: Debrand-Passard et al. (Eds.), Synthèse géologique du Sud-Est de la France, Mémoire BRGM, France 125, pp. 263–272.
Farinacci, A. (2002). Western Tethys Jurassic gaps as windows on the land. Interaction between hot-spots and gaps. Palaeopelagos Special Publication 4, 32 p.
Farinacci, A., Bölükbasi, S., & Ridolfi, V. (1997). The Tethyan Jurassic “main gap” in the Tinaz Tepe section of the Barla Dag area, Western Taurus, Turkey. Palaeopelagos, 7, 17–26.
Fuchs, W. (1973). Ein Beitrag zur Kenntnis der Jura “Globigerinen” und verwandter Formen an Hand polnischen Materials des Callovien and Oxfordien. Verhandlungen der Geologische Bundesanstalt, 3, 445–487.
Fülöp, J. (1975). The Mesozoic Basement Horst Blocks of Tata. (Tatai mezozoós alaphegység rögök.). Geologica Hungarica, series Geologia 16, 225 p.
Gaillard, C. (1983). Les Biohermes à Spongiaires et leur Environnement dans l’Oxfordien du Jura Méridional. Documents des Laboratoires de Géologie, Université Claude-Bernard, Lyon 90, 515 p.
Gawlik, H.-J., & Schlagintweit, F. (2006). Berriasian drowning of the Plassen carbonate platform at the type-locality and its bearing on the early Eoalpine orogenic dynamics in the Northern Calcareous Alps (Austria). International Journal of Earth Sciences Geologische Rundschau, 95, 451–462.
Gianotti, A. (1958). Deux facies du Jurassique supérieur en Sicile. Revue de Micropaléontologie, 1, 38–51.
Giovagnoli, M. C. & Schiavinotto, F. (1986). Indagini biometriche su foraminiferi globigeriniformi del Dogger-Malm umbro-marchigiano: indicazioni preliminari. In: Piccinini, R. (Ed.), Atti convegno “Fossili Evoluzione Ambiante”, Pergola, 1984, pp. 131–134.
Giovagnoli, M. C., & Schiavinotto, F. (1987). Dati morphometrici su foraminiferi globigeriniformi del Bajociano-Kimmeridgiano di Colle Tordina (Appennino umbro-marchigiano). Bollettino della Società Paleontologica Italiana, 25, 187–197.
Giovagnoli, M. C., & Schiavinotto, F. (1990). Definizione biometrica dell’ontogenesi di globigeriniformi nel Jurassico umbro-marchigiano (Appennino centrale). Geologica romana, 26, 109–125.
Gorbachik, T. N., & Kuznetsova, K. I. (1983). Jurassic and Early Cretaceous planktonic foraminifera (Favusellidae). Stratigraphy and paleobiogeography. Zitteliana, 10, 459–466.
Gorbatchik, T. N. (1986). Yurskie i rannemelovie planktonnie foraminiferi yuga SSSR. Jurassic and Early Cretaceous planktonic foraminifera from the South SSSR. Akademia Nauk SSSR, 238 p.
Gorbatchik, T. N., & Antonova, Z. A. (1981). Revision of some species of planktonic Lower Cretaceous foraminifers in the connection with the study them in the electron microscope. Voprosy mikropaleontologii, Akademia Nauk, SSSR, 24, 82–87.
Gorbatchik, T. N., & Kuznetsova, K. I. (1998). Problems on the systematics and nomenclature of the Early Planktonic Foraminifers. Paleontological Journal, 32(3–11), 549–558.
Görög, Á., & Wernli, R. (2002). The Middle and Late Bathonian protoglobigerinids of Gyenespuszta (Bakony Mts, Hungary). Revue de Paléobiologie, 20, 12–23.
Görög, Á., & Wernli, R. (2003). Palaeobiogeography of the Middle Jurassic protoglobigerinids (Foraminifera). Eclogae geologicae Helvetiae, 96, 237–249.
Görög, Á., & Wernli, R. (2004). A rare protoglobigerinid association (Foraminifera) from the Tithonian of Gerecse Mts, Hungary. Hantkeniana, 4, 37–45.
Gradstein, F. M. (1976) Biostratigraphy and biogeography of Jurassic Grand Banks foraminifera. In: Schafer, C.T. & Pelletier, B.R (Eds.), First international symposium on benthonic foraminifera of continental margins. Part B, paleoecology and biostratigraphy. Maritime sediments, Special Publication 1, pp. 557–583.
Gradstein, F. M. (1983). Paleoecology and stratigraphy of Jurassic abyssal foraminifera in the Blake-Bahama basin, Deep Sea drilling project site 534. In: Sheridan et al. (Eds.), Initial reports of the Deep Sea drilling project 86, pp. 537–559.
Grigelis, A. A. (1958). Globigerina oxfordiana n.sp., an occurence of Globigerina in the Upper Jurassic strata of Lithuania: Nauchnye Doklady Vysshei Shkoly. Geologo-Geographicheskie Nauki, 3, 109–111.
Grigelis, A. A. & Gorbatchik, T. N. (1980a) The systematics of Jurassic and Early Cretaceous Globigerinacea. Paleontolgical Journal, 6–17.
Grigelis, A., & Gorbatchik, T. (1980b). Morphology and taxonomy of Jurassic and Early Cretaceous representatives of the superfamily Globigerinacea (Favusellidae). Journal of Foraminiferal Research, 10, 180–190.
Grigelis, A. A., Mesezhnikov, M. S., Yakovleva, S. P., & Kozlova, G. E. (1977). The first finds of planktonic foraminifera in the Upper Jurassic of the Pechora river basin. Doklady Akad. Nauk SSSR, 233, 926–927.
Grigelis, A. A. & Norling, E. (1999) Jurassic geology and foraminiferal faunas in the NW part of the East European Platform. A Lithuanian -Swedish geotraverse study. Research Papers, SGU series Ca 89, Forskningsrapporter, 101 p.
Gušić, J., & Babić, L. (1970). Some biostratigraphic and lithogenetic characteristics of the Jurassic in Zumberak (Northwest Croatia). Geološki Vjesnik, 23, 39–54.
Haas, J., Jocha-Edelényi, E., Gidai, L., Kaiser, M., Kreitzoi, M. & Oravecz, J. (1984) Geology of the surroundings of Sümeg. Geologica Hungarica, Series Geologica 20, 365 p.
Haeusler, R. (1881). Note sur une zone à Globigerines dans les terrains Jurassiques de la Suisse. Annales de la Société Royale Malacologique de Belgique, 16, 188–190.
Heinz, A. R., & Isenschmid, Chr. (1988). Mikrofazielle und stratigraphische Untersuchungen im Massivkalk (Malm) der Préalpes médianes. Eclogae geologicae Helvetiae, 81, 1–62.
Hudson, W., Hart, M. B., Sidorczuk, M. & Wierzbowski, A. (2005). Jurassic planktonic foraminifera from Pieniny Klippen Belt and their taxonomic and phylogenetic importance (Carpathians, southern Poland). Volumina Jurassica (Tomy Jurajskie) III, pp. 1–10.
Iovčeva, P. & Trifonova, E. (1961). Tithonian Globigerina from North-West Bulgaria. Travaux sur la Géologie de Bulgarie, série. Paléontologie, Académie des Sciences Bulgarie 3, pp. 343–351.
Krajewski, M., & Olszewska, B. (2007). Foraminifera from the Late Jurassic and Early Cretaceous carbonate platform facies of the southern part of the Crimea Mountains; Southern Ukraine. Annales Societatis Geologorum Poloniae, 77, 291–311.
Kuznetsova, K. I. & Gorbatchik, T. N. (1985). Upper Jurassic and Lower Cretaceous stratigraphy and foraminifers of the crimea. Nauka, Moskow: Academy of Sciences SSSR, 136, 218 p.
Kuznetsova, K. I., Grigelis, A. A., Adjamian, J., Jamarkini, E. & Hallaq, L. (1996). Zonal stratigraphy and foraminifera of the Tethyan Jurassic (Eastern Mediterranean). Amsterdam: Gordon & Breach Publishers, 256 p.
Kuznetsova, K. I. & Seibold, I. (1978). Foraminifers from the Upper Jurassic and Lower Cretaceous of the Eastern Atlantic (DSDP Leg. 41, sites 367 and 370). In: Lancelot, Y. et al. (Eds.), Reports of the Deep Sea drilling project 41, pp 515–537.
Kuznetsova, K. I. & Uspenskaya, E. A. (1980). Novie nakhdki planktonnikh foraminifer v verkhneyurskikh otlozhenii Krima. (New finds of planktonic foraminifera in the Upper Jurassic deposits of the Crimea). Doklady Akademii Nauk SSSR (Reports of the Academy of Sciences of the USSR, Earth Science Sections) 254, 242–246 (748–752).
Langoisseux, O. (2001). Stratigraphie séquentielle basée sur une approche multidisciplinaire: Le Kimméridgien inferieur de la Fosse Vocontienne (S-E France). Unpublished Master thesis, University of Geneva, 89 p.
Leischner, W. (1961). Zur Kenntnis der Mikrofauna und -flora der Salzburger Kalkalpen. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 112, 1–47.
Lethiers, F., & Crasquin-Soleau, S. (1988). Comment extraire les microfossiles à tests calcitiques des roches calcaires dures. Revue de Micropaléontologie, 31, 56–61.
Luterbacher, H. (1972). Foraminifera from the Lower Cretaceous and Upper Jurassic of the northwestern Atlantic. Initial Reports of the Deep Sea drilling project, 11, 561–593.
Masters, B. A. (1977). Mesozoic planktonic foraminifera. In: Ramsay, A. T. S. (Ed.), Oceanic micropaleontology 1. London: Academic Press, pp. 301–731.
Mišik, M. (1959). “Lombardiová” mikrofácia vedúci horizont v malme západnych Karpát. (Die “Lombardia”- Mikrofazies–ein Leithorizont im Malm, der Westkarpaten.) Geologicky Sbornik Slovenska Akademia Ved. X, pp. 171–181.
Morozova, V. G., & Moskalenko, T. A. (1961). Foraminiferes planctoniques des dépôts limitrophes du Bajocien et du Bathonien du Daghestan central (NE du Caucase). Voprosy Micropaleontologii SSSR, 5, 3–30.
Nebert, K. (1964). Zur geologie des Kelkit Cay-Oberlaufs Südwestlich von Siran (Nordostanatolien). Mineral Research and Exploration Institute of Turkey, pp. 42–59.
Oesterle, H. (1968). Foraminiferen der Typlokalität der Birmenstorfer-Schischten, unterer Malm (Teilrevision der Arbeiten von J. Kübler & H. Zwingli 1866–1870 und von R. Haeusler 1881–1883). Eclogae geologicae Helvetiae 61, 695–792.
Papp, A. & Turnovsky, K. (1970). Anleitung zur biostratigraphischen Auswertung von Gesteinsschliffen (Microfacies Austriaca). Jahrbuch der Geologischen Bundesanstalt, Sonderband 16, 50 p.
Pazdrowa, O. (1969). Bathonian Globigerina of Poland. Annales de la société Geologique de Pologne, 39, 41–56.
Perconig, E. (1962). Sur la constitution géologique de l’Andalusie occidentale en particulier du Bassin de Guadalquivir (Espagne méridonal). Livre à Mémoire du Prof. Paul Fallot, 1: Mémoires de la Société Géologique de France, pp. 229–256.
Riegraf, W. (1987a). Planktonic foraminifera (Globuligerinidae) from the Callovian (Middle Jurassic) of Southwest Germany. Journal of Foraminiferal Research, 17, 190–211.
Riegraf, W. (1987b). Planktonische Foraminiferen und Radiolarien im Callovium und Oxfordium (Jura) Süddeutschlands. Neues Jahrbuch für Geologie Paläntologie—Abhandlungen, 176, 91–103.
Riegraf, W. (1988). Callovian (Middle Jurassic) benthonic foraminifera from southwest Germany. Revista Española de Micropaleontología, 20, 451–490.
Riegraf, W., & Luterbacher, H. (1989). Oberjura-Foraminiferen aus dem Nord-und Südatlantik (Deep Sea drilling project Leg 1–79). Geologische Rundschau, 78, 999–1045.
Rod, E. (1937). Stratigraphie des Malm der Graustock-Hutstock-Gruppe (Melchtal, Kanton Obwalden). Bern: Stämpfli & Cie, 55 p.
Samson, Y. (1997). Utilisation des Foraminiféres dans l’ estimation des variations bathymétriques des environnements de dépôt marins Jurassiques: Applications au Kimméridgien de l’ Ouest-Européen. Mémoires des sciences de la Terre de l’Université Pierre & Marie Curie (Paris VI) 97(10), 394 p.
Samson, Y. (2001). Foraminiferes et reconstitution des variations bathymétriques: example du Kimméridgien de la région du Havre (Seine-Maritime, Normandie, France). Revue de Micropaléontologie, 44, 59–91.
Simmons, M. D., BouDagher-Fadel M. K., Banner F. T. & Whittaker, J. E. (1997). The Jurassic Favusellacea, the earliest Globigerinina. In: BouDagher-Fadel et al. (Eds.), The early evolutionary history of planktonic foraminifera. London: Chapmann & Hall, pp. 17–51.
Stam, B. (1986). Quantitative analysis of Middle and Late Jurassic foraminifera from Portugal and its implications for the Grand Banks of Newfoundland. Utrecht Micropaleontological Bulletins 34, 168 p.
Subbotina, N. N. (1953). Globigerinidae, Hantkeninidae and Globorotaliidae. In: Proceedings of the All-Union Oil Research Geological Survey Institute, new series 76, 296 p.
Terquem, M. O. (1883). Cinquième mémoire sur les foraminifères du système oolithique de la zone à Ammonites parkinsoni de Fontoy (Moselle). Bulletin de la Société géologique de France, 11, 339–406.
Thierry, J. (2000). Early Kimmeridgian. In: Dercourt et al. (Eds.), Atlas Peri-Tethys. Paleogeographical maps-explanatory notes. CCGM/CGMW, Paris, map 10, 85–97.
Wernli, R. (1988). Les protoglobigérines (foraminiferes) du Toarcien et de l’Aalénien du Domuz Dag (Taurus occidental, Turquie). Eclogae geologicae Helvetiae, 81, 661–668.
Wernli, R., Ascoli, P. & Williams, G.L. (1995). Favusella hoterivica (Subbotina) from the Berriasian and Valanginian of offshore Eastern Canada. Revue de Paléobiologie 14, 379–398.
Wernli, R., & Görög, Á. (1999). Protoglobigerinids (Foraminifera) acid extracted from Bajocian Limestones (Hungary). Revista Española de Micropaleontología, 31, 123–130.
Wernli, R., & Görög, Á. (2000). Determination of Bajocian protoglobigerinids (Foraminifera) in thin sections. Revue de Paléobiologie, 19, 399–407.
Wernli, R., & Görög, Á. (2007). Protoglobigérines et Oberhauserellidae (Foraminifères) du Bajocien-Bathonien du Jura méridional, France. Revue de Micropaléontologie, 50, 185–205.
Wernli, R. & Kindler, P. (1986). Les “Protoglobigérines” du Callovo-Oxfordien de Châtillon-sur-Cluses (Préalpes internes, Haute-Savoie, France). Eclogae geologicae Helvetiae 79, 137–147.
Acknowledgments
Sincere thanks are due to F. Athrops (Lyon) for his information about the Crussol field section. We benefited from the assistance of P. Desjacques (Labo processing), R. Martini (SEM photography), G. Frébourg (thin sections photography) and J. Metzger (infography) who are warmly thanked. Thank to J.-M. Jaquet who greatly improved the English. We are grateful to the referees M. BouDagher-Fadel and to M. B. Hart for their comments and suggestions and to S. Spezzaferri and D. Marty for their exhaustive editorial work. The fieldwork has been supported by the “Fondation Dr. Joachim De Giacomi” of the Swiss Academy of Sciences and by the Hungarian OTKA Foundation (contract K68791), which is greatly acknowledged.
Author information
Authors and Affiliations
Corresponding author
Additional information
Editorial handling: Silvia Spezzaferri & Daniel Marty.
Rights and permissions
About this article
Cite this article
Görög, Á., Wernli, R. Kimmeridgian protoglobigerinids (Foraminifera) from Crussol (SE France). Swiss J Geosci 103, 83–100 (2010). https://doi.org/10.1007/s00015-010-0010-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00015-010-0010-8