- Published:
The Pliensbachian ammonites of the Algarve Basin (Portugal) and their palaeobiogeographical significance for the “Iberia-Newfoundland” conjugate margins
Swiss Journal of Geosciences volume 104, pages 81–96 (2011)
Abstract
A review of the Pliensbachian ammonite faunas of the Algarve Basin is given covering their taxonomy, stratigraphy and palaeobiogeography (palaeobiodiversity). This review is based on both an extensive revision of the previously collected and/or published material and on new finds precisely located within the ammonite succession. This new material allows us to clarify the variations within the most abundant species—a new species Reynesocoeloceras elmii nov. sp. is described—and to improve our understanding (palaeobiodiversity, palaeobiogeography and bio- or chrono-stratigraphy) of the two single known Pliensbachian fossiliferous assemblages in the Algarve Basin. The first of these assemblages is relatively diverse and is ascribed to the upper half of the Luridum Subchronozone. The second assemblage is less diversified and probably partly condensed. It is broadly attributed to the upper half of the Stokesi Subchronozone. Despite the newly collected ammonites and extensive prospecting, our knowledge of the Pliensbachian ammonite faunas of the Algarve remains fragmentary. Therefore, it is difficult to propose an incontrovertible extensive palaeobiogeographical interpretation for these faunas, but it seems that Tethyan (Mediterranean) affinities were of major importance whereas there were probably no NW European influences via the Lusitanian Basin.
1 Introduction
The Algarve Basin was one of the numerous pre-drift basins of the “Iberia-Newfoundland” conjugate margins. Most of these basins—which first formed during the Early Mesozoic—lie offshore and we generally know nothing of their Jurassic faunas (or at least their macrofaunas). All of the basins located on the North American margin in the vicinity of Newfoundland are situated offshore and only the Lusitanian Basin and to a lesser extent the Algarve Basin, which lie on the western and south-western Iberian margin respectively, have significant Early Jurassic onshore outcrops. With an unusually large proportion of endemic taxa during the Late Sinemurian, the Early Pliensbachian and to a lesser extent during the Late Pliensbachian (Dommergues 1987; Dommergues and Mouterde 1987; Dommergues and El Hariri 2002; Dommergues et al. 2004, 2010; Mouterde et al. 2007) the rich and well-known ammonite faunas of the Lusitanian Basin are palaeontologically distinct from those of the NW European and Mediterranean areas as a whole. The intriguing, highly endemic Lusitanian faunas raise the question of the palaeobiogeographical significance of the “Iberia-Newfoundland” conjugate margins during the Early Jurassic. In addition to the unusual number of endemic taxa, the Lusitanian faunas are also remarkable because they are clearly associated with the NW European province and not with the Tethyan realm, although all the available palaeogeographical reconstructions suggest a clear structural opening of the Lusitanian areas towards the Mediterranean basins of the western Tethys (Dommergues and El Hariri 2002; Dommergues et al. 2004, 2010). Accordingly, any new information about the Early Jurassic ammonite faunas of any basins located in the vicinity of the Lusitanian Basin may be of major importance to our understanding of the puzzling palaeobiogeography of the “Iberia-Newfoundland” conjugate margins. The present study of the Pliensbachian faunas of the Algarve Basin enhances that understanding.
While Middle and Late Jurassic ammonites are rather common in the Algarve Basin, Early Jurassic and especially Pliensbachian ones are much less abundant and are very restricted both spatially and stratigraphically. However, rare and incomplete as they are, these faunas provide the only possible comparisons with the Lusitanian faunas from anywhere on the “Iberia-Newfoundland” conjugate margins. The present study is based on most of the material previously collected in the Algarve by P. Choffat (collection of the “Serviços Geológicos de Portugal”) and by R.B. Rocha (collections of the “Universidade Nova de Lisboa”)—some of these specimens were illustrated by Rocha (1971a, b)—and on the quite numerous ammonites collected recently within a precise stratigraphic framework and housed in the collections of the “Université de Bourgogne”. The present work seeks firstly to provide a synthesis of the Pliensbachian ammonite faunas of the Algarve Basin based on a full taxonomic revision of all the available ammonites, and secondly to propose palaeogeographical and palaeoecological interpretations of these faunas in the context of the pre-drift basins of the “Iberia-Newfoundland” conjugate margins.
2 Geographical and geological setting
An extensive historical synthesis of the geological and palaeontological explorations of the Algarve Basin is given by Rocha (1977: 17–25). This synthesis is used here as reference material and is not repeated. Among the numerous works cited by Rocha (1977) few are dedicated to the stratigraphy and/or palaeontology of the Early Jurassic (Choffat 1887; Pratsch 1958; Rocha 1971a, b; Rocha et al. 1979, 1983). In fact, except for a short preliminary publication (Rocha 1971b), the monograph on the Jurassic of the western Algarve Basin by Rocha (1977) is the first and the only significant palaeontological work. Rocha (1977) provides accurate descriptions and some photographic illustrations of ammonites properly situated in the Early Jurassic successions of the Algarve Basin. Since that work, only a few studies of the Early Jurassic of the Algarve Basin have been published and those are mainly about its structural and/or sedimentological features (e.g., Terrinha et al. 2002; Ribeiro and Terrinha 2007). The present work is the first palaeontological publication about the Pliensbachian of the Algarve since Rocha’s monograph (1977).
Rocha (1977) reports only two fossiliferous localities with Pliensbachian ammonites for the whole of the Algarve Basin (Fig. 1). These two localities are coastal outcrops (cliffs and/or rocky slopes) between Sagres and Cabo de S. Vicente (Fig. 2). The first of these fossiliferous localities is restricted to a few scattered exposures of beds 492–496 in the thick series of cherty limestones cropping out close to Cabo de S. Vicente (Figs. 2, 3, 4, 5) (cf. Rocha 1977: 31). The second locality is also very limited, being confined to the top of a single bed (conventionally numbered 1000) of “crystalline compact limestones with Fuciniceras” (sensu Rocha 1977: 69) cropping out close to the top of the near vertical coastal cliffs, about 400 m east of Forte de Belixe (Figs. 2, 3, 4, 5, 6) (cf. Rocha 1977: 31–33). At this spot the cliffs are mostly a thick series (about 50–55 m) of cherty limestones overlain by about 1.5 m of “crystalline limestones with Brachiopods” followed by about 4 m of “crystalline compact limestones with Fuciniceras”. Unfortunately, the outcrops are discontinuous close to the top of the cliffs and it is difficult to specify the exact location of fossiliferous bed 1000. Nevertheless, it is probably situated a little above the top of the “crystalline limestones with Brachiopods” and we estimate that the two fossiliferous episodes with Pliensbachian ammonites of the Algarve Basin lie several tens of metres apart.

Sketch map showing the location and extent of the main Jurassic outcrops in Portugal. The outcrops of the Lusitanian and Algarve Basins are indicated by striped and shaded areas, respectively

Sketch map showing the location of the studied fossiliferous localities (A–C) on or close to the costal cliffs between Sagres and Cabo de S. Vicente (Sagres, Algarve, Portugal)

Sketch map showing the location of the fossiliferous outcrops on the costal exposures to the East (B 1, B 2) and Northeast (A) of Cabo de S. Vicente (Sagres, Algarve, Portugal)

Photograph of the fossiliferous outcrops (B 1, B 2) on the costal exposures East of Cabo de S. Vicente (Sagres, Algarve, Portugal). The location of the vantage point is indicated in Fig. 3

Lithological pattern (erosion) and fossiliferous beds with ammonites in the Pliensbachian (cherty limestones) section East of Cabo de S. Vicente (Sagres, Algarve, Portugal). The number of specimens collected is given for each taxon and each level

Sketch map showing the location of the fossiliferous outcrop (C) situated close to the top of the near vertical coastal cliffs about 400 m East of Forte de Belixe (Sagres, Algarve, Portugal)
Despite recent systematic prospecting of the Early Jurassic series cropping out between Sagres and Cabo de S. Vicente, no new fossiliferous locality with Pliensbachian ammonites has been found. As a result our knowledge of the Pliensbachian faunas of the Algarve remains very fragmentary. However, the new finds, housed in the collections of the “Université de Bourgogne”, do provide a better understanding of variability within several species [e.g., Reynesocoeloceras elmii nov. sp., Fuciniceras gr. marianii (Fucini, 1904) and Fuciniceras gr. celebratum (Fucini, 1900)] and also a better understanding of the biodiversity of the two faunal assemblages. As a result, and with just a single fossiliferous episode for the Early Pliensbachian and another one for the base of the Late Pliensbachian, comparisons with other basins (e.g., the Lusitanian Basin) are possible but any generalization is difficult and partly speculative.
Palaeogeographically, the Algarve Basin is one of the numerous basins of the “Iberia-Newfoundland” conjugate margins of the Atlantic Ocean. More specifically, like the Porto, Lusitanian and Alentejo Basins, it is one of the inner basins of the Iberian margin (Fig. 7). Among all the basins of the “Iberia-Newfoundland” conjugate margins, the Lusitanian and Algarve Basins are the only ones with onshore fossiliferous Pliensbachian outcrops and the only ones to yield Pliensbachian ammonites. In addition, the Algarve Basin occupies an interesting palaeobiogeographic position in the south-western part of the Iberian margin, close to the westernmost confines of the Tethyan realm.

Pre-drift palaeogeographical sketch map of the “Iberia-Newfoundland” conjugate margins with location of the main basins. The Iberian landmass is outlined in its present-day configuration as a landmark. The location and extent of Mesozoic pre-drift basins (shaded regions) are only approximate. Modified from Srivastava and Verhoef (1992) and Sandness and Pacheco (2002). The 2,000 m isobath is suggested by dotted lines to approximate the offshore outlines of the Newfoundland and Iberian landmasses. Plate boundaries cannot be pinpointed
3 Systematic palaeontology
- Class:
-
Cephalopoda Cuvier, 1797
- Subclass:
-
Ammonoidea Zittel, 1884
3.1 Phylloceratoidea
- Suborder:
-
Phylloceratina Arkel, 1950
- Superfamily:
-
Phylloceratoidea Zittel, 1884
- Family:
-
Juraphyllitidae Arkell, 1950
- Genus:
-
Juraphyllites Müller, 1939
- Type species:
-
Phylloceras diopsis Gemmellaro, 1884, by original designation
3.1.1 Juraphyllites aff. libertus (Gemmellaro, 1884) (Fig. 8l, m)
aff. 1884 Phylloceras libertum Gemmellaro, pl. 2, fig. 1-5.

Early Pliensbachian (Ibex Chronozone, Luridum Subchronozone) ammonites from the cherty limestones of Cabo de S. Vicente (Sagres, Algarve, Portugal). a–k Reynesocoeloceras elmii nov. sp., level 496. a Holotype, CSV.02.01 (This specimen was previously published by Rocha (1977: pl. 1, fig. 5.5). b–k Paratypes. b UBGD 277367. c UBGD 277368. d UBGD 277369. e UBGD 277370. f SGP.A.26/G.B6 (this specimen was previously published by Rocha (1977: pl. 1, fig. 5.6). g UBGD 277371. h UBGD 277372. i UBGD 277373. j UBGD 277374. k UBGD 277375. l, m Juraphyllites aff. libertus (Gemmellaro, 1884), level 496. l UBGD 277376. m UBGD 277377. n Juraphyllites aff. helveticus Wiedenmayer, 1977, level 496, UBGD 277378. o Lytoceras gr. fimbriatum (J. Sowerby, 1817), level 492, UBGD 277379. Specimens a–n are coated with magnesium oxide, specimen o with ammonium chloride. Scales bar 10 mm (a–n), 20 mm (o)
Remarks
Although fragmentary and partly corroded these two small, silicified specimens can be credibly attributed to the genus Juraphyllites and more precisely to the group of J. libertus taken in a broad sense. The rounded ribs visible on the upper flank and chiefly on the ventral area where they project in a forward-directed chevron are a possible diagnostic feature for J. libertus. Nevertheless, this attribution must remain tentative because (1) the subelliptical whorl cross-section is appreciably broader than that of the lectotype illustrated by Gemmellaro (1884: pl. 2, fig. 1-3) and designated by Fantini-Sestini (1974) and (2) no constrictions are visible on the fragmentary Algarve specimens. If one ignores the absence of constrictions, the specimen may be compared, although not very convincingly, with the Sicilian exemplar illustrated and designated as “Phylloceras mimatensis d’Orbigny” by Gemmellaro (1882: pl. 12, fig. 24).
Stratigraphic and geographic ranges
In the Algarve, J. aff. libertus is located in bed 496 (top) of the Cabo de S. Vicente section. The age of this bed is roughly the upper half of the Luridum Subchronozone. J. libertus is a usually common, chiefly Mediterranean taxon and is known for example in Italy (Apennines, Sicily, Southern Alps), Austria and Germany (Austoalpine units), Switzerland (Southern Alps), Hungary (Bakony), Morocco (Middle and High Atlas, Beni-Snassen), Tunisia (Dorsale), Portugal (Algarve), Slovakia (Fatra), Turkey (Pontids) and Georgia (Caucasus). In the vicinity of the Mediterranean areas, the species is also known from a few specimens collected in some southern localities of NW Europe, e.g. France (Causses, Subbriançonnais), Switzerland (Helvetic Basin) and Romania (Carpathians). The long stratigraphic range of J. libertus begins close to the base of the Early Pliensbachian (Jamesoni Chronozone) and ends in the Toarcian. The presence of Juraphyllites aff. libertus in the Algarve is not very significant for bio- and chrono-stratigraphy, but it is very informative palaeobiogeographically. The species J. libertus and more generally the genus Juraphyllites are unknown in the widely documented faunas of the Lusitanian Basin and their occurrence in the Algarve clearly suggests Mediterranean affinities.
3.1.2 Juraphyllites aff. helveticus Wiedenmayer, 1977 (Fig. 8n)
aff. 1977 Juraphyllites helveticus Wiedenmayer, pl. 18, fig. 15-16.
Remarks
Although it is partly crushed and corroded this small incomplete silicified Juraphyllites exhibits a distinctive ornamental pattern. While the subplatycone shell morphology with its subelliptical whorl cross-section is commonplace for the genus and is similar to that of J. libertus for example, the precocious coarse ribbing is unusual. At similar diameters the inner whorls of most Juraphyllites are ribless and, except for some possible constrictions, totally smooth. By contrast, the specimen illustrated in Fig. 8n has prominent ribbing from about 2 cm in diameter. The ribs start close to mid-flank, become clearly prorsiradiate on the ventral area and cross the venter in a curved chevron. The ribs reach their greatest height on the ventral area. The single roughly comparable form illustrated in the literature is Juraphyllites helveticus, a possibly Late Sinemurian and/or Early Pliensbachian species collected in the Tessin (Switzerland, Southern Alps) and known only by the holotype. The imperfect state of preservation of the Swiss and Portuguese specimens precludes any unambiguous attribution. The inner whorls of the Swiss ammonite (before about 3 cm in diameter) are corroded and the two forms cannot be compared at exactly the same diameters. Despite a possible ontogenetic bias, the ribbing pattern of the Swiss specimen looks coarser than that of the Portuguese ammonite. The two forms cannot unreservedly be paired.
Stratigraphic and geographic ranges
In the Algarve, J. aff. helveticus is located in bed 496 (top) of the Cabo de S. Vicente section. The age of this bed is roughly the upper half of the Luridum Subchronozone. Juraphyllites helveticus is a rare Mediterranean taxon until now known in the Southern Alps (Tessin, Switzerland) only. The existence of a rather similar form in the Algarve Basin is consistent with the Mediterranean affinities of the Algarve Pliensbachian ammonite faunas. Conversely this form is poorly informative in stratigraphic terms and at best suggests the Late Sinemurian and/or Early Pliensbachian.
3.2 Lytoceratoidea
- Suborder:
-
Psiloceratida Housa, 1965
- Superfamily:
-
Lytoceratoidea Neumayr, 1875
- Family:
-
Lytoceratidae Neumayr, 1875
- Subfamily:
-
Lytoceratinae Neumayr, 1875
- Genus:
-
Lytoceras Suess, 1865
- Type species:
-
Ammonites fimbriatus J. Sowerby, 1817, by original designation
3.2.1 Lytoceras gr. fimbriatum (J. Sowerby, 1817) (Fig. 8o)
1817 Ammonites fimbriatus J. Sowerby, pl. 164.
Remarks
This specimen is an incomplete and more or less crushed cast of light beige limestone. It has a very ordinary shell morphology and ornamentation for the genus Lytoceras. The whorl cross-section is somewhat crushed but clearly rounded (probably subcircular or broadly subelliptic before deformation). The whorls expand quickly but with little overlap and the umbilicus is rather wide [umbilical width index (UWI) ≈ 0.40]. The very dense ribbing pattern consists of annular, radial, thin and often crenulate riblets. They do not usually bifurcate but may sometimes be almost jointed close to the umbilical seam. All these traits clearly designate the Portuguese ammonite as a member of the group of Lytoceras fimbriatum.
Stratigraphic and geographic ranges
In the Algarve, Lytoceras gr. fimbriatum is located in bed 492 (top) of the Cabo de S. Vicente section. The age of this bed is roughly the upper half of the Luridum Subchronozone. Lytoceras gr. fimbriatum is an often common, widespread (almost ubiquitous) and long-ranged taxon, which is poorly informative for both palaeobiogeography and stratigraphy. The presence of this species in the Early Pliensbachian of the Algarve Basin is not surprising but does not allow us to clarify the paleobiogeographical affinities of the faunas.
3.3 Eoderoceratoidea
- Suborder:
-
Psiloceratida Housa, 1965
- Superfamily:
-
Eoderoceratoidea Spath, 1929
- Family:
-
Dubariceratidae Dommergues & Meister, 1999
- Subfamily:
-
Reynesocoeloceratinae Dommergues, 1986 (emended Dommergues and Meister, 1999)
- Genus:
-
Reynesocoeloceras Géczy, 1976
- Type species:
-
Coeloceras crassum Young & Bird var. indunensis Meneghini, 1881, by original designation
3.3.1 Reynesocoeloceras elmii n. sp. (Fig. 8a–k)
-
1887 cf. Aegoceras densinodum (Quenstedt) in Wright—Choffat, p. 235 (=Fig. 8f).
-
1971b Metaderoceras sp.—Rocha, pp. 170-174, pl. 1, fig. 1, 2 (=Fig. 8f).
-
1977 Platypleuroceras sp.—Rocha, p. 96, pl. 1, fig. 5.5 (=Fig. 8a).
-
1977 Metaderoceras sp. nov.—Rocha, pp. 94–96. pl. 1, fig. 5.6 (=Fig. 8f).
-
1983 Reynesocoeloceras praeincertum Dommergues & Mouterde—Rocha et al., p. 16.
Type material
An incomplete, rather large, but mostly well-preserved silicified specimen is selected as the holotype (CSV.02.01). This holotype was first illustrated by Rocha (1977, pl. 1, fig. 5.5) as Platypleuroceras sp. It is illustrated again (after complete acid dissolution of the carbonated gangue) in this work (Fig. 8a). The suture lines are missing but if allowance is made for the ontogenetic variations in the ornamental pattern, one can suspect that the body chamber is, at least partly, preserved. Ten additional, more or less incompletely preserved specimens are designated as paratypes (Fig. 8b–k) (UBGD 277367, UBGD 277368, UBGD 277369, UBGD 277370, SGP.A.26/G.B6, UBGD 277371, UBGD 277372, UBGD 277373, UBGD 277374, UBGD 277375). One of these paratypes (SGP.A.26/G.B6) (Fig. 8f) first illustrated by Rocha (1971b, pl. 1, fig. 1, 2) as “Metaderoceras sp.” belongs to the collection of the “Geological Survey of Portugal”. It was labelled by hand and first cited by Choffat (1887, p. 235) as “cf. Aegoceras densinodum (Quenstedt) in Wright” and comes from Cabo de S. Vicente. The paratypes illustrate the ontogeny and/or the intraspecific variation of the new species.
Type horizon and locality
Outcrops of cherty limestones, approximately 100 m east of the lighthouse of Cabo de S. Vicente, bed 496 (upper part). Sagres, Algarve, Portugal.
Derivation of the name
In honour of Serge Elmi for his extensive knowledge of the Jurassic in general and of the Portuguese Jurassic ammonites in particular.
Material
In addition to the holotype and the 10 paratypes, 11 other often crushed, incomplete and somewhat silicified Reynesocoeloceras were collected from bed 496 of the Cabo de S. Vicente section. These specimens are not illustrated but can be convincingly attributed to the new species. They are housed in the collections of the “Université de Bourgogne”. Suture lines are missing but some alterations of the ornamental pattern suggest that the body chamber is often, at least partly, preserved.
Diagnosis
Medium-sized Reynesocoeloceras with—but for precocious fine secondary and intercalary ribs on the ventral area—a rather short (about 2.5 cm diameter) Metaderoceras-type juvenile-stage. This first growth stage is usually quickly superseded by one that persists until the end of the body chamber. This late stage of ontogeny is marked by plainly evolute coiling—the shell becomes subserpenticone—with barely arched subparallel flanks and an ornamental pattern of crowded subannular ribs invariably bearing tiny but sharp prominent ventrolateral tubercles as far as the end of the body chamber. The ribs are interrupted at the ventrolateral edge by a tubercle. In fact, it is the combination of (1) a subquadratic section (with an almost flat ventral area) and of (2) the persistence (even in the final growth stages) of tiny sharp ventrolateral tubercles that are the best diagnostic traits of the new species.
Description
The adult diameter of this medium-sized Reynesocoeloceras probably rarely exceeds 7 cm. A size of 5–6 cm is frequent. Possible dimorphism (microconch vs. macroconch) cannot be documented from the available material. Unambiguous signs of maturity are missing but shell shape and ornamental pattern clearly change during ontogeny. The last whorl of the large specimens (probably mainly the body chamber) and the inner-medium whorls must therefore be described separately. In the inner and medium whorls, the shell is subplatycone to subplanorbicone with a usually slightly to faintly depressed subcadicone (Fig. 8f, j) to confusedly rectangular (Fig. 8e) whorl cross-section. The slightly arched to almost flat flanks usually diverge gently from the faint umbilical shoulders towards the clearly angular ventrolateral margins. The depressed low ventrolateral area is evenly arched but usually slightly convex (Fig. 8f). The straight and more or less prorsiradiate ribs arise in the umbilical area. They grow more prominent on the sides and culminate in a ventolateral tubercle or spine (Fig. 8a, b, e–k). Beyond this point the ribs usually subdivide into (usually two or three) regularly spaced secondary ribs projecting gently forwards towards the venter (Fig. 8d, f, h–k). In the inner and medium whorls the ribbing density and prominence are rather variable from widely spaced and coarse (Fig. 8d, f) to closely spaced and fine (Fig. 8a, b, i, j). To sum up, in the inner and medium stages of growth, the habitus of the new species is, with the exception of the obvious ventral ribbing, rather similar to the Metaderoceras-style.
In the later stages of growth (suspected end of the phragmocone and/or body chambers), the shell habitus clearly changes and intraspecific variability declines. The shell is subserpenticone with a slight overlap of the previous whorl. The whorl cross-section is slightly compressed and subrectangular in outline with subparallel high flanks and a gently arched depressed wide ventral area. The greatest whorl width occurs at about the first third of whorl height. The umbilicus is shallow. The umbilical margin is gently rounded and the boundary between the umbilical area and the flank is just perceptible. Conversely, and especially as regards ornamentation, the ventrolateral margin is always distinct and often quite prominent, probably as far as the end of the body chamber. The ornamentation consists of crowded ribbing (Fig. 8a, b, g). While the juvenile ribbing is coarse, the transition to the late dense ribbing is usually rapid (Fig. 8d, f). The moderately high prorsiradiate simple ribs begin close to the umbilical seam. At the ventrolateral margin, the ribs rise gently and bear tiny but distinct and sharp, radially elongate tubercles (Fig. 8a, g). These tubercles probably persist until the end of the body chamber. Unlike on the inner whorls, the ventral ribs are not subdivided and do not fade beyond the ventrolateral margins in the late stages of growth.
Discussion
With more or less clear Metaderoceras-style juvenile stages, the new species is similar to Reynesocoeloceras praeincertum Dommergues & Mouterde, 1982, a Mediterranean and Lusitanian taxon that can be interpreted as a transitional form between the genera Metaderoceras and Reynesocoeloceras (Dommergues & Mouterde 1982; Dommergues 1987, 1990). The new species is probably a part of this important but complex phylogenetic transition, which is chiefly controlled by various peramorphic processes affecting (at different rates?) shell morphology, ornamentation and the suture line. Moreover, several close, but separate evolutionary branches can be suspected (Dommergues 1987). Thus, in Reynesocoeloceras elmii n. sp., (1) the Metaderoceras-style juvenile stages are usually shorter and not as distinctive as in R. praeincertum, (2) the morphological and ornamental pattern of the late stages of growth are transitional between those of R. praeincertum—which bear fairly strong ventrolateral tubercles till the end of the body chamber—and those of the more derived and younger species, as for example R. gr. indunense (Meneghini, 1981)—fallax (Fucini, 1905)—which have crowded annular (without tubercles) ribs on the body chamber.
In fact, in R. elmii n. sp., while ribs are often crowded and almost annular on the body chamber, they are invariably interrupted by a tiny but well defined, sharp ventrolateral tubercle (Fig. 8a, b, g). Somewhat similar characteristics can also be observed in R. incertum (Fucini, 1905), but in this Mediterranean species the ornamental pattern is coarser and less variable during growth. The marked contrast between the juvenile and late stages of ontogeny, which is usually observed in R. elmii n. sp., seems to be missing in R. incertum.
The new Portuguese species can also be compared with some North American forms, Reynesocoeloceras grahami Smith & Tipper, 1996, R. cf. incertum sensu Smith and Tipper (1996) non Fucini (1905) and Reynesocoeloceras sp. sensu Johannson et al. (1997). However, the highly evolute coiling—a trait which is visible from the inner whorls—and consequently the quasi-serpenticone shells of these North American ammonites exclude any phyletic proximity between them and the Mediterranean and/or Portuguese forms. A convergence phenomenon may be assumed.
Stratigraphic and geographic ranges
In the Algarve, Reynesocoeloceras elmii n. sp. is located in bed 496 (top) of the Cabo de S. Vicente section. The age of this bed is roughly the upper half of the Luridum Subchronozone. With the exception of some North American forms, the genus Reynesocoeloceras is clearly a Mediterranean taxon, chiefly known in Italy (Apennines, Southern Alps), Austria (Austoalpine units), Switzerland (Southern Alps), Hungary (Bakony), Portugal (Algarve) and Morocco (High Atlas). Some species, usually only represented by rare specimens, are also known from a few southern localities of NW Europe, as for example in France (Causses, Lyon area) but also in the Portuguese Lusitanian Basin where R. praeincertum is common but restricted to a single bed of the Crassum zonule in the Peniche section (Dommergues and Mouterde 1982; Dommergues 1987).
Until now, the new species Reynesocoeloceras elmii n. sp. has only been found in bed 496 of the Cabo de S. Vicente section in the Algarve, a locality where it is accompanied by taxa that are all poorly informative in stratigraphic terms: Juraphyllites aff. libertus, Juraphyllites aff. helveticus and Lytoceras gr. fimbriatum. Owing to the lack of possible correlations, the age of the new species can only be assumed by indirect arguments based on the evolutionary pattern of the genus Reynesocoeloceras. An age subsequent to the praeincertum horizon is credible and the fossiliferous deposit of Cabo de S. Vicente probably corresponds approximately to the late part of the Luridum Subchronozone.
3.4 Hildoceratoidea
- Suborder:
-
Psiloceratida Housa, 1965
- Superfamily:
-
Hildoceratoidea Hyatt, 1867
- Family:
-
Hildoceratidae Hyatt, 1867
- Subfamily:
-
Harpoceratinae Neumayr, 1875
- Genus:
-
Fuciniceras Haas, 1913
- Type species:
-
Hildoceras lavinianum Meneghiniin Fucini1900, by original designation
N.B.: As previously proposed by Dommergues et al. (2002: 459–462), we use the name Fuciniceras in a broad meaning including all the Pliensbachian Harpoceratinae usually designated in the literature as Fuciniceras Haas, 1913 or Protogrammoceras Spath, 1913. For example, in a recent review of the Late Pliensbachian ammonites (Mouterde et al. 2007), the names Fuciniceras or Protogrammoceras are used in a traditional way. Nevertheless, and taking into account the state of the art, these two genera in their common uses are no more than polyphyletic groups based on still poorly understood homoplasies (e.g., convergence, parallelism and/or reversion).
3.4.1 Fucinicerascelebratum (Fucini, 1900) (Figs. 9a, c, g, 10e)
-
1900 Grammoceras celebratum Fucini, p. 41, pl. 10, fig. 1-2.
-
1977 Protogrammoceras celebratum (Fucini, 1900)—Rocha, pp. 105–106, pl. 1, fig. 5.8 (=Fig. 9c).
Fig. 9 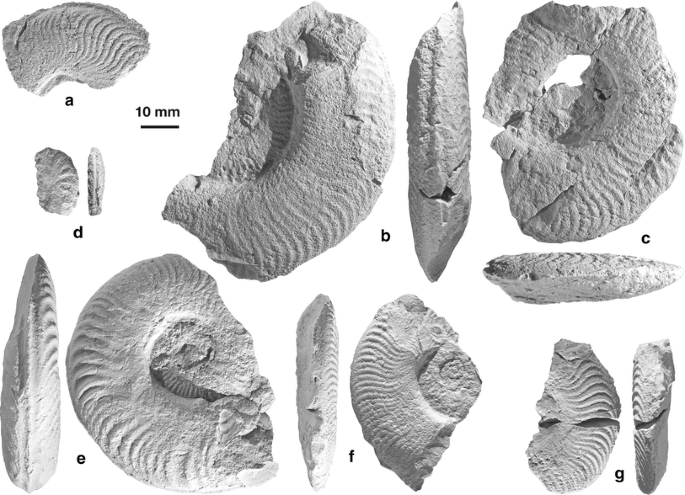
Late Pliensbachian (Margariatus Chronozone, Stokesi Subchronozone) ammonites from the “crystalline compact limestones with Fuciniceras” (sensu Rocha 1977: 69), East of Forte de Belixe, level 1000 (Sagres, Algarve, Portugal). a, c, g Fuciniceras celebratum (Fucini, 1900), a UBGD 277380. c BEL.08.10 (this specimen was previously published by Rocha (1977: pl. 1, fig. 5.8). g UBGD 277382. b, f Fuciniceras aff. marianii (Fucini, 1900), b UBGD 277381. f BEL.08.05 (this specimen was previously published by Rocha (1977: pl. 1, fig. 5.7). e Fuciniceras marianii (Fucini, 1904), BEL.08.33 (this specimen was previously published by Rocha (1977: pl. 1, fig. 5.9). d Arieticeras sp., UBGD 277391. All the specimens are coated with magnesium oxide. Scale bar 10 mm

Description
These more or less incomplete specimens are casts made of compact light beige limestone. They are slightly crushed and distorted but the main features can be easily and confidently observed. Suture lines are not usually preserved. The shells of these medium- to moderate-sized Fuciniceras are intermediate between the platycone and oxycone types with an obviously compressed elliptical whorl section and a fairly wide umbilicus (UWI ≈ 0.35). The greatest whorl width is between mid-flank and the umbilical margin. The umbilicus is rounded with no angular shoulder. The transition between the umbilical wall and the moderately to barely arched base of the flanks is distinct but gradual. Conversely, the transition between the flanks and the ventral area is faint with no perceptible ventrolateral margin. The upper flanks noticeably and consistently converge towards the narrow venter, which bears an obvious sharp keel. The ribs are closely spaced and sinuous. At least on the casts, these clearly falcoid ribs begin on the lowermost flank and end at the base or very close to the base of the keel. Usually the keel is not flanked by smooth bands. If present, these bands are very narrow, clearly sloped and never prominent.
Discussion
The specimens studied here are very similar to the Italian lectotype illustrated by Fucini (1900) and designated by Wiedenmayer (1977). In this sense the taxonomic attribution is clear. Nevertheless, and partly because the Portuguese ammonites are included in a probably fairly condensed assemblage, it is not possible to base the attribution on an unbiased appraisal of intraspecific variations. For example, the occurrence of transitional forms between Fuciniceras celebratum and F. marianii within the condensed assemblage diminishes the contrasts between species.
Stratigraphic and geographic ranges
In the Algarve, Fuciniceras celebratum is located in bed 1000 (top) of a section, which crops out about 400 m east of Forte de Belixe. The age of this somewhat condensed bed belonging to the “crystalline compact limestones with Fuciniceras” (sensu Rocha 1977: 69) is roughly the upper half of the Stokesi Subchronozone. Fuciniceras celebratum is chiefly a Mediterranean taxon which is known for example in Italy (Apennines, Calabria, Sicily, Southern Alps), Austria and Germany (Austoalpine units), Switzerland (Southern Alps), Hungary (Bakony), Morocco (Middle and High Atlas, Beni-Snassen), Tunisia (Dorsale), Spain (Betic ranges), Portugal (Algarve) and Slovakia (Fatra). Outside of the Mediterranean areas the species is also known in some southern localities of NW Europe, e.g. Spain (Iberic ranges), France (Causses, Provence, Corbières, Lyon area) and the Lusitanian Basin (Mouterde et al. 2007). In the southern part of NW Europe and in the Lusitanian Basin F. celebratum occurs in the Celebratum zonule which is the last subdivision of the Stokesi Subchronozone. In the Mediterranean areas, the range of F. celebratum probably begins slightly earlier (i.e., as early as the Nitescens zonule) but in any case, F. celebratum does not reach the Subnososus subchronozone. In this context, and as suggested by the possible transitional form between F. celebratum and F. marianii, one can suspect that the range of Fuciniceras celebratum in the Algarve is the same as in the Mediterranean areas (i.e., part of the Nitescens zonule and Celebratum zonule).
3.4.2 Fuciniceras cf. celebratum (Fucini, 1900) (Fig. 10c, h)
cf. 1900 Grammoceras celebratum Fucini, p. 41, pl. 10, fig. 1-2.

Late Pliensbachian (Margariatus Chronozone, Stokesi Subchronozone) ammonites from the “crystalline compact limestones with Fuciniceras” (sensu Rocha 1977: 69), east of Forte de Belixe, level 1000 (Sagres, Algarve, Portugal). a, b Fuciniceras marianii (Fucini, 1904). a UBGD 277386. Arrow indicates the beginning of the body chamber. b UBGD 277387. c, h Fuciniceras cf. celebratum (Fucini, 1900). c UBGD 277384. h UBGD 277385. d Fuciniceras aff. marianii (Fucini, 1900), UBGD 277388. e Fuciniceras celebratum (Fucini, 1900), UBGD 277383. f, g Fuciniceras cf. marianii (Fucini, 1900). f UBGD 277389. g UBGD 277390. All the specimens are coated with magnesium oxide. Scale bar 10 mm
Remarks
These two specimens are too incompletely preserved to allow an unreserved assignment to Fucini’s species—the ventral area is for example difficult to observe—but the coiling and ribbing patterns well match those of Fuciniceras celebratum.
3.4.3 Fucinicerasmarianii (Fucini, 1904) (Figs. 9e, 10a, b)
-
1904 Harpoceras marianii Fucini, p. 283, pl. 41, fig. 1-3.
-
1977 Protogrammoceras exiguum (Fucini, 1904)—Rocha, pp. 104–105, pl. 1, fig. 5.9 (=Fig. 9e).
Description
These ammonites are incomplete and/or slightly crushed and distorted casts of light beige limestone. Suture lines are barely preserved. Shells are subplatycone with a compressed whorl section and a somewhat open umbilicus (UWI ≈ 0.34). The whorl section is subogival. Up to the upper two-thirds of whorl height, the flanks are hardly arched and subparallel. But they curve and converge towards the venter on the outer third. The venter bears a sharp keel, which is flanked by two flat or slightly depressed and narrow but apparently smooth bands. The umbilical margin is distinct but rounded with no angular shoulder. The rather dense ribbing is formed by falcoid to subfalcoid ribs. They arise on the lower flank and end just before reaching the smooth ventral bands. The peculiar ribless lower flank of the specimen illustrated in Fig. 9e is perhaps due to trauma.
Discussion
If compared with all the Pliensbachian Fuciniceras s.l., F. marianii and F. celebratum are obviously two very similar species. They are consecutive taxa in the Mediterranean fossil record (e.g., Mouterde and Ferretti 1986; Geczy and Meister 1998) and it is tempting to consider F. marianii as a possible close ancestor of F. celebratum. Thus, Mouterde and Ferretti (1986) propose a phyletic Mediterranean succession leading from Fuciniceras isseli (Fucini, 1900) to F. celebratum via F. marianii. In this context, transitional steps between these three species can be suspected. Compared with F. celebratum, the main distinctive features of F. marianii are (1) the less clearly ogival whorl section with subparallel flanks up to the upper two thirds of whorl height and the rather rounded but not tapered ventral area and (2) the existence of always perceptible, more or less sloping, flat bands flanking the keel. The rib outline is subfalcoid or falcoid in F. marianii but always falcoid in F. celebratum. Moreover, it is difficult to distinguish between F. marianii and F. bonarellii (Fucini, 1900)—at least in the sense of Faraoni et al. (2002)—two probably partly coeval forms. F. bonarellii typically has a more depressed and flatter ventral area and so a subogival to subrectangular whorl section (Faraoni et al. 2002). Another confusing species is Fuciniceras exiguum (Fucini, 1904), especially if the small and poorly preserved lectotype illustrated by Fucini (1904: pl. 19, fig. 8) and designated by Wiedenmayer (1977) is taken as a reference. It is difficult to state whether this form belongs to the group of Fuciniceras volubile (Fucini, 1900) [including F. pantanelli (Fucini, 1900) and F. varicostatum (Fucini, 1900)] or if it is close to the group of F. marianii–F. celebratum. In his previous study of the Algarve ammonite faunas Rocha (1977) attributed the specimen illustrated in Fig. 9e to Fuciniceras exiguum. In the present work we prefer to interpret this form as an atypical (perhaps with some traumatic marks) specimen of F. marianii. In any case, not all of the specimens from the Algarve studied and illustrated here can be unreservedly attributed to F. marianii. They are perhaps transitional towards F. celebratum or representative of a slightly diverging group.
Stratigraphic and geographic ranges
In the Algarve, Fuciniceras marianii is located in bed 1000 (top) of the section, which crops out about 400 m east of Forte de Belixe. The age of this somewhat condensed bed, which belongs to the “crystalline compact limestones with Fuciniceras” (sensu Rocha 1977: 69), is roughly the upper half of the Stokesi Subchronozone.
F. marianii is a chiefly Mediterranean taxon known in Italy (Apennines, Southern Alps), Austria (Austroalpine units), Hungary (Bakony) and in Morocco (High Atlas). The species is also known in some rare southern localities of NW Europe such as France (Causses, Provence) and Switzerland (Helvetic Basin). The short range of this species characterizes a horizon in the middle part of the Mediterranean lavinianum biozone. This Mediterranean horizon is probably roughly equivalent to the Nitescens standard zonule.
3.4.4 Fuciniceras aff. marianii (Fucini, 1904) (Figs. 9b, f, 10d)
aff. 1904 Harpoceras marianii Fucini, p. 283, pl. 41, fig. 1-3.
Remarks
These incompletely preserved ammonites probably belong to the group of Fuciniceras marianii if it is considered in a broad sense. They have subfalcoid ribs are perceptibly less flexuous (especially the specimen illustrated in Fig. 9f), more slender and/or crowded than is usual for F. marianii. Moreover, the umbilicus of the forms illustrated in Fig. 9b, f looks rather wide for the group. In view of such traits, the specimen illustrated in Fig. 9f was previously attributed by Rocha (1977) to Fuciniceras isseli. In the present work we prefer a tentative attribution to the group of F. marianii s.l. but with some reserve and considering the possibility of various degrees of transition between F. isseli and F. marianii. Nevertheless, taking into account (1) the imperfect preservation of the specimens and (2) the condensation of the fossiliferous deposit and consequently the absence of stratigraphic constraints, it is difficult to reach any firm conclusion.
Stratigraphic and geographic ranges
In the Algarve, Fuciniceras aff. marianii is located in bed 1000 (top) of the section cropping out about 400 m east of Forte de Belixe. The age of this somewhat condensed bed is roughly the upper half of the Stokesi Subchronozone. If the Portuguese ammonites designated here as Fuciniceras aff. marianii are transitional forms between F. isseli and F. marianii, one can suspect an age in the late part and/or the early part of the isseli and marianii Mediterranean horizons respectively.
3.4.5 Fuciniceras cf. marianii (Fucini, 1904) (Fig. 10f, g)
cf. 1904 Harpoceras marianii Fucini, p. 283, pl. 41, fig. 1-3.
Remarks
These two partly corroded and more or less incomplete casts are tentatively assigned to the group of F. marianii s.l. The specimen illustrated in Fig. 10f is a nucleus. It exhibits the rounded aspect of the ventral area and the rather blunt keel typical of the juvenile stages of the species.
3.4.6 Arieticeras sp. (Fig. 9d)
- Subfamily:
-
Arieticeratinae Howarth, 1955
- Genus:
-
Arieticeras Seguenza, 1885
- Type species:
-
Ammonites algovianus Oppel, 1862, by original designation
Remarks
This small specimen is only a piece of a whorl of a rather evolute (possibly subplatycone) shell. It has spaced but moderately coarse ribbing. The flanks are gently arched and the ventral area bears a keel flanked by narrow (apparently depressed?) smooth bands. All of these traits suggest an Arieticeratinae or even an Arieticeras. The hypothesis of a coarsely ribbed and evolute Harpoceratinae (e.g., Fieldingiceras) is possible but less convincing, so the assumption of an Arieticeras is preferred here. In stratigraphic terms this form is interesting because the Arieticeratinae family is alleged to be absent from the Stokesi Subchronozone and usually the very first Arieticeras does not appear in the fossil record before the base of the subsequent Subnodosus Subchronozone. In the Algarve stratigraphic context, the presence of this possible Arieticeras is surprising because all the other ammonites collected in the fossiliferous locality of Forte de Belixe clearly indicate the Stokesi Subchronozone. Two possibilities can be considered: (1) the condensed interval of Forte de Belixe encompasses the base of the Subnodosus Subchronozone, (2) the small collected specimen is an exceptionally precocious member of the Arieticeratinae family.
4 Paleobiogeographical and stratigraphic implications
Early Jurassic deposits in the Algarve older than the Toarcian contain few if any ammonite fossils. The Sinemurian is marked by a 60 m thick, chiefly dolomitic, unfossiliferous series. These “Dolomite and dolomite limestones of Espiche” (sensu Rocha 1977: p. 69) are overlain by several tens of metres of often cherty bedded limestones roughly corresponding to the Pliensbachian. Within all these Pliensbachian limestones only two brief fossiliferous episodes with ammonites are known (Fig. 11) (Rocha 1977).

Stratigraphic framework of the fossiliferous episodes with Pliensbachian ammonites in the Algarve Basin. The ages of the fossiliferous episodes are compared with the standard chronostratigraphic scale (chronozones, subchronozones and zonules) and with two biostratigraphic scales: (1) the Lusitanian scale of horizons and/or bioevents and (2) a Mediterranean scale (biozones and horizons and/or bioevents). Conventionally and in contrast to the names of the standard chronostratigraphic units, those of the biostratigraphic units are italicized
The first episode, which is documented in beds 492 and chiefly 496 of Cabo da S. Vicente (Fig. 5), has quite numerous benthic (bivalves, gastropods, brachiopods) and pelagic (ammonites, belemnites) fossils. With its four species from three separate superfamilies and two separate suborders, the ammonite fauna of this first fossiliferous episode is well diversified in taxonomic terms (even if species richness is unexceptional). However, it is difficult to propose an indisputable bio or chronostratigraphic hypothesis because the taxa are either endemic (i.e., Reynesocoeloceras elmii nov. sp.), long-ranged (i.e., Juraphyllites aff. libertus, Lytoceras gr. fimbriatum), or poorly documented in terms of bio- or chrono-stratigraphy (i.e., J. aff. helveticus). Nevertheless, indirect arguments based on what is known about the evolutionary history of the genera Metaderoceras and Reynesocoeloceras allow us to suggest an age roughly corresponding to the later part of the Luridum Subchronozone for Reynesocoeloceras elmii nov. sp. (Fig. 11). In palaeobiogeographical terms and if one excludes Lytoceras gr. fimbriatum, which is a widespread species fairly common both in the NW European and Mediterranean areas, all the other taxa belonging to the genera Reynesocoeloceras and Juraphyllites suggest clear Tethyan affinities. However, because at the species level R. elmii nov. sp. is probably an endemic taxon, the Algarve Basin may have been partly isolated during the Ibex Chronozone (Luridum Subchronozone).
The second fossiliferous episode is clearly distinct from the first one both in stratigraphic and palaeoecological terms. It is known from a single locality about 400 m east of Forte de Belixe where a small and isolated fossiliferous outcrop of “crystalline compact limestones with Fuciniceras” (sensu Rocha 1977: p. 69) is situated close to the top of the cliffs. The ammonites are rather abundant but strictly confined to this small outcrop, which is the upper part of a single bed. This second fossiliferous outcrop lies several tens of metres (probably about 40 m) above the first one.
Except for a possible poorly-preserved Arieticeras, all the ammonites collected from the east of Forte de Belixe belong to a single genus (i.e., Fuciniceras). The faunal assemblage suggests a possible minor condensation in the Margaritatus Chronozone. Indeed the occurrence of species such as Fuciniceras celebratum and F. marianii indicates that at least two successive Mediterranean biohorizons are probably involved (Fig. 11). These ammonites are found together with belemnites, bivalves and brachiopods. Nevertheless, if one allows for the probable condensation of the deposit, the ammonite fauna is poorly diversified and monotonous in taxonomic terms. The absence of Phylloceratoidea and Lytoceratoidea must be underlined.
The Pliensbachian ammonite faunas of the Algarve are obviously very partially known because only two brief and separate episodes are documented. Nevertheless the available data convincingly suggest Tethyan (Mediterranean) affinities. Indeed, if one excepts the poorly informative Lytoceras gr. fimbriatum which is, at least from the Ibex Chronozone onwards, a virtually ubiquitous taxon, and Fuciniceras celebratum which is a common Mediterranean form but which is also known in several southern localities of the NW European province [e.g., Spain (Iberic ranges) France (Causses, Provence, Corbières, Lyon area) and Portugal (Lusitanian Basin)], all the other species collected in the Algarve are reliable indicators of Mediterranean faunas. Even the new species Reynesocoeloceras elmii nov. sp., which is probably an endemic form, belongs to a genus that is primarily known in the Mediterranean areas [e.g., Italy (Apennines, Southern Alps), Austria (Austroalpine units), Switzerland (Southern Alps), Hungary (Bakony), Portugal (Algarve) and Morocco (High Atlas)]. In fact, within the Portuguese paleobiogeographical framework, there is no evidence of free faunal exchanges between the Algarve and Lusitanian Basins. As far as the Pliensbachian and more specifically the Early Pliensbachian ammonite faunas are concerned, Portugal seems to be a palaeogeographically heterogeneous area. Indeed, chiefly during the Early Pliensbachian, the Lusitanian faunas are clearly dominated by NW European faunal influences (Dommergues and Mouterde 1987; Dommergues and El Hariri 2002; Dommergues et al. 2004, 2010), while these are unknown in the Algarve Basin. By extension, and although only the Luridum and Stokesi Subchronozones are partially documented, the Algarve Basin may have been palaeobiogeographically a Mediterranean entity free from NW European faunal influences during the Pliensbachian. This tentative proposal differs from the hypothesis proposed by Terrinha et al. (2002: fig. 8), which supposes there were NW European faunal influences in the Algarve Basin from the Late Sinemurian onwards and assumes that the Lusitanian and Algarve Basins had similar palaeontological histories. In fact, despite their geographical proximity on the western and south-western margin of Iberia, the two basins seem hardly connected and their faunal (ammonite) compositions are not readily comparable. This palaeobiogeographical phenomenon is surprising and difficult to explain.
However, belemnite rostra are fairly common in the Pliensbachian fossiliferous succession of the Algarve: a phenomenon that is usual in the NW European province but quite uncommon in the Mediterranean basins. This suggests a complex palaeobiogeographical setting for the Algarve Basin.
5 Conclusions
Despite recent prospecting, only the two Pliensbachian fossiliferous localities previously recognized in the Algarve and described by Rocha (1971a, b, 1977) (i.e., Cabo de S. Vicente and east of Forte de Belixe) have been found again. With the exception of these two outstanding stratigraphic levels, the Pliensbachian of the Algarve seems to be devoid of ammonites. As a result, our knowledge of the Pliensbachian ammonite faunas of the Algarve remains very discontinuous and incomplete with a first fossiliferous episode during the Luridum Subchronozone in the Early Pliensbachian and a second one during the Stokesi Subchronozone in the Late Pliensbachian. Nevertheless, new findings have significantly improved our taxonomic understanding of the previously collected forms—most importantly a new species, Reynesocoeloceras elmii, is described—and greatly enhanced our knowledge of palaeobiodiversity. For example, the discovery in the Algarve of ammonites belonging to the Juraphyllitidae during the Luridum Subchronozone changes our view of the palaeobiogeographic affinities of the Algarve faunas. The new data suggest close affinities with the Tethyan realm (i.e., the Mediterranean province) and not, as previously suspected (Terrinha et al. 2002), with the Lusitanian Basin and by extension with the NW European Province. In fact, although relatively close to one another, the Algarve and Lusitanian Basins seem to be related to two separate palaeobiogeographical realms and it is not possible to consider the “Iberia-Newfoundland” conjugate margins, in general, and the western and south-western Iberian margins in particular as homogenous palaeobiogeographical entities. This result raises the question of the existence of possible structural and/or ecological barriers within the “Iberia-Newfoundland” conjugate margins and leaves the problem of Lusitanian endemism unresolved.
Abbreviations
- BEL:
-
Forte de Belixe, specimen housed in the collections of the “Universidade Nova de Lisboa” (R.B. Rocha collection)
- CSV:
-
Cabo de Sao Vicente, specimen housed in the collections of the “Universidade Nova de Lisboa” (R.B. Rocha collection)
- SGP:
-
Specimen housed in the Collections of the “Serviços Geológicos de Portugal” (P. Choffat collection)
- UBGD:
-
Specimen housed in the collections of the “Université de Bourgogne, Géologie Dijon”
References
Choffat, P. (1887). Recherches sur les terrains secondaires au Sud du Sado. Communicações da Commissao dos Trabalhos Geologicos, 1, 222–311.
Dommergues, J.-L. (1987). L’évolution chez les Ammonitina du Lias moyen (Carixien, Domérien basal) en Europe occidentale. Documents des Laboratoires de Géologie Lyon, 98, 1–297.
Dommergues, J.-L. (1990). Ammonoids. In K. J. McNamara (Ed.), Evolutionary trends (pp. 162–187). London: Belhaven Press.
Dommergues, J.-L., & El Hariri, K. (2002). Endemism as a palaeobiogeographic parameter of basin history illustrated by early- and mid-Liassic peri-Tethyan ammonite faunas. Palaeogeography, Palaeoclimatology, Palaeoecology, 184, 407–418.
Dommergues, J.-L., & Meister, C. (1999). Cladistic formalisation of relationships within a superfamily of lower Jurassic Ammonitina: Eoderocerataceae Spath, 1929. Revue de Paléobiologie, 18, 273–286.
Dommergues, J.-L., Meister, C., & Mouterde, R. (2002). Fuciniceras paradoxus nov. sp. (Harpoceratinae, Ammonitina) du Domérien portugais. Réflexion sur le sens taxonomique d’un assemblage paradoxal de caractères. Geobios, 35, 457–468.
Dommergues, J.-L., Meister, C., Neige, P., & Rocha, R. B. (2004). Endemic Sinemurian (Early Jurassic) ammonites from the Lusitanian Basin (Portugal). Revue de Paléobiologie, 23, 529–549.
Dommergues, J.-L., Meister, C., & Rocha, R. B. (2010). The Sinemurian ammonites of the Lusitanian Basin (Portugal): a remarkable example of complex endemic evolution. Palaeodiversity, 3, 139–167.
Dommergues, J.-L., & Mouterde, R. (1982). Reynesocoeloceras praeincertum nov. sp. (Ammonitina, Carixien moyen). Remarques sur l’origine du genre Reynesocoeloceras. Geobios, 15, 757–763.
Dommergues, J.-L., & Mouterde, R. (1987). The endemic trends of the Liassic ammonite faunas of Portugal as the result of the opening up of a narrow epicontinental basin. Palaeogeography, Palaeoclimatology, Palaeoecology, 58, 129–137.
Fantini-Sestini, N. (1974). Phylloceratina (Ammonoidea) del Pliensbachiano italiano. Rivista Italiana di Paleontologia, 80, 193–250.
Faraoni, P., Marini, A., Pallini, G., & Venturi, F. (2002). Protogrammoceratinae and new ammonite assemblages of the central Apennines and their significance on the Carixian-Domerian biostratigraphic boundary in the Mediterranean paleoprovince. Geologica Romana, 36, 215–249.
Fucini, A. (1900). Ammoniti del Lias Medio dell’Appennino Centrale existensi nel museo di Pisa. Palaeontographia italica, 6, 17–78.
Fucini, A. (1904). Cefalopodi Liassici del Monte di Cetona. Palaeontographia Italica, 10, 241–264.
Fucini, A. (1905). Cefalopodi Liassici del Monte di Cetona. Palaeontographia Italica, 11, 265–318.
Geczy, B., & Meister, C. (1998). Les ammonites du Domérien de la montagne du Bakony (Hongrie). Revue de Paléobiologie, 17, 69–161.
Gemmellaro, G. G. (1882). Sopra alcune faune giuresi e liasiche della Sicilia. Palermo: Stabilimento tipographico.
Gemmellaro, G. G. (1884). Sui fossili degli strati a Terebratura Aspasia della contrada Rocche rosse presso Galati (Provincia di Messina). Giornale di Scienze naturali ed economiche Palermo, 16, 167–218.
Johannson, G. G., Smith, P. L., & Gordey, S. P. (1997). Early Jurassic evolution of the northern Stikinian arc: Evidence from the Laberge Group, northwestern British Columbia. Canadian Journal of Earth Sciences, 34, 1030–1057.
Mouterde, R., Dommergues, J.-L., Meister, C., & Rocha, R. B. (2007). Atlas des fossiles caractéristiques du Lias portugais. III a. Domérien (Ammonites). Ciências da Terra, 16, 67–111.
Mouterde, R., & Ferretti, A. (1986). Les Ammonites—Caractères et intérêt. Exemple de l’évolution des Harpocératidés du Lias moyen mésogéen (Apennin des Marches). In G. Pallini (Ed.), Atti I convegno Pergola 1984: Fossili, Evoluzione, Ambiente (pp. 73–77). Pergola: Comitato Centenario Raffaele Piccinini.
Pratsch, J. C. (1958). Stratigraphisch-tektonische Untersuchungen im Mesozoikum von Algarve (Südportugal). Beihefte zum Geologischen Jahrbuch, 30, 1–123.
Ribeiro, C., & Terrinha, P. (2007). Formation, deformation and chertification of systematic clastic dykes in differentially lithified carbonate multilayer. SW Iberia, Algarve Basin, Lower Jurassic. Sedimentary Geology, 196, 201–215.
Rocha, R. B. (1971a). Contribuição para o estudo geológico da região do Cabo de S. Vicente-Alvor. Boletim da Sociedade Geológica de Portugal, 17, 139–168.
Rocha, R. B. (1971b). Un Metaderoceras (Carixien) du Cap S. Vicente (Algarve). Boletim da Sociedade Geológica de Portugal, 17, 169–176.
Rocha, R. B. (1977). Estudo estratigráfico e paleontológico do Jurássico do Algarve ocidental. Ciências da Terra, 2, 1–178.
Rocha, R. B., Ramalho, M. M., Antunes, M. T., & Coelho, A. V. P. (1983). Carta Geológica de Portugal na escala de 1/50000. Notíca explicative da folha 52-A Portimão. Serviços Geológicos de Portugal.
Rocha, R. B., Ramalho, M. M., Manuppella, G., Zbyszewski, G., & Coelho, A. V. P. (1979). Carta Geológica de Portugal na escala de 1/50000. Notíca explicative da folha 51-B Vila do Bispo. Serviços Geológicos de Portugal.
Sandness, F., & Pacheco, J. (2002). Portuguese deepwater basins show potential. Offshore, 62, 1–3.
Smith, P. L., & Tipper, H. W. (1996). Pliensbachian (Lower Jurassic) Ammonites of the Queen Charlotte Islands, British Columbia. Bulletins of American Paleontology, 108, 1–122.
Sowerby, J. (1812–1822). The mineral conchology of Great Britain (Vol. 1–4 (début), pl. 1–383). London: Benjamin Meredith.
Srivastava, S. P., & Verhoef, J. (1992). Evolution of Mesozoic sedimentary basins around the North Central Atlantic: a preliminary plate kinematic solution. In J. Parnell (Ed.), Basins of the Atlantic Seaboard: Petroleum geology, sedimentology and basin evolution. Geological Society (London) Special Publication, 62, pp. 397–420.
Terrinha, P., Ribeiro, C., Kullberg, J. C., Lopes, C., Rocha, R., & Ribeiro, A. (2002). Compressive episodes and faunal isolation during rifting, Southwest Iberia. The Journal of Geology, 110, 101–113.
Wiedenmayer, F. (1977). Die Ammoniten des Besazio-kalks (Pliensbachian, Südtessin). Mémoires suisses de Paléontologie, 98, 1–169.
Acknowledgments
This paper is a contribution by the FED “Forme, Evolution, Diversité” team of the “Biogéosciences” research unit (UMR 5561, CNRS/uB) and under the FCT/MCT–POCTI/36438/PAL/2000 BIOSCALES Program. We thank Christopher Sutcliffe for help with the English version. We are grateful to Cyril Hugues Dommergues for his fine photographic work. The authors are grateful to Alain Morard for his constructive review of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Editorial handling: Hugo Bucher & Daniel Marty.
Rights and permissions
About this article
Cite this article
Dommergues, JL., Meister, C. & Rocha, R.B. The Pliensbachian ammonites of the Algarve Basin (Portugal) and their palaeobiogeographical significance for the “Iberia-Newfoundland” conjugate margins. Swiss J Geosci 104, 81–96 (2011). https://doi.org/10.1007/s00015-011-0056-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00015-011-0056-2